Addison, P.F.E., D.J. Collins, R. Trebilco, S. Howe, N. Bax, and P. Hedge. 2018. A new wave of marine evidence-based management: Emerging challenges and solutions to transform monitoring, evaluating, and reporting. ICES Journal of Marine Science 75(3):941–952, https://doi.org/10.1093/icesjms/fsx216.
Allison, G.W., S.D. Gaines, J. Lubchenco, and H.P. Possingham. 2003. Ensuring persistence of marine reserves: Catastrophes require adopting an insurance factor. Ecological Applications 13:S8–S24, https://doi.org/10.1890/1051-0761(2003)013[0008:EPOMRC]2.0.CO;2.
Babcock, R.C., N.T. Shears, A.C. Alcala, N.S. Barrett, G.J. Edgar, K.D. Lafferty, T.R. McClanahan, and G.R. Russ. 2010. Decadal trends in marine reserves reveal differential rates of change in direct and indirect effects. Proceedings of the National Academy of Sciences of the United States of America 107(43):18,256–18,261, https://doi.org/10.1073/pnas.0908012107.
Babcock, E.A., and A.D. MacCall. 2011. How useful is the ratio of fish density outside versus inside no-take marine reserves as a metric for fishery management control rules? Canadian Journal of Fisheries and Aquatic Sciences 68(2):343–359, https://doi.org/10.1139/F10-146.
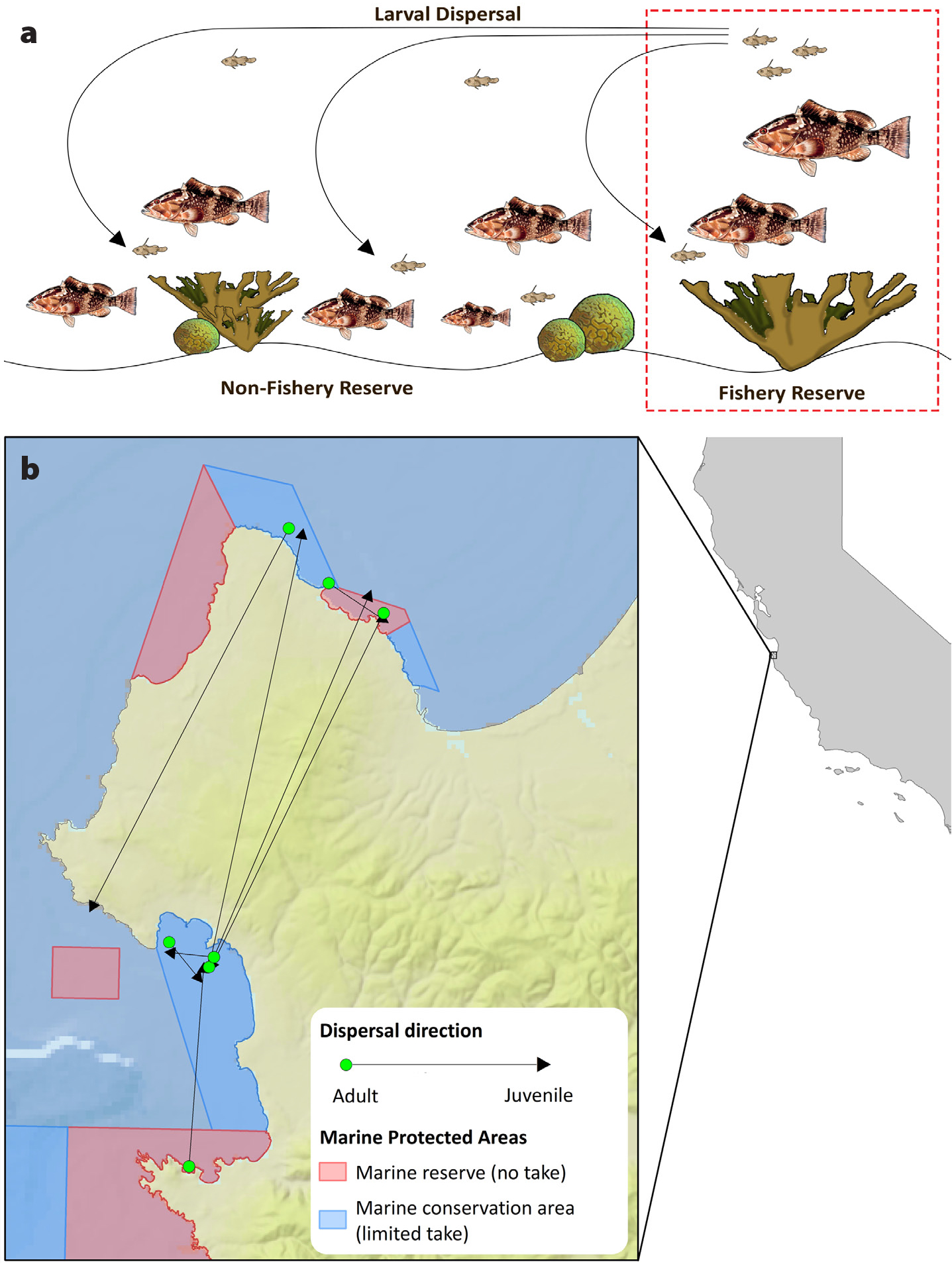
Baetscher, D.S., E.C. Anderson, E.A. Gilbert-Horvath, D.P. Malone, E.T. Saarman, M.H. Carr, and J.C. Garza. 2019. Dispersal of a nearshore marine fish connects marine reserves and adjacent fished areas along an open coast. Molecular Ecology 28(7):1,611–1,623, https://doi.org/10.1111/mec.15044.
Ball, I.R., and H.P. Possingham. 2001. The design of marine protected areas: Adapting terrestrial techniques. Pp. 769–774 in Proceedings of the International Congress on Modelling and Simulation. F. Ghassemi, P. Whetton, R. Little, and M. Littleboy, eds, The Modelling and Simulation Society of Australia and New Zealand Inc.
Barneche, D.R., D.R. Robertson, C.R. White, and D.J. Marshall. 2018. Fish reproductive-energy output increases disproportionately with body size. Science 360(6389):642–645, https://doi.org/10.1126/science.aao6868.
Baskett, M.L., and L.A.K.K. Barnett. 2015. The ecological and evolutionary consequences of marine reserves. Annual Review of Ecology, Evolution, and Systematics 46(1):49–73, https://doi.org/10.1146/annurev-ecolsys-112414-054424.
Bates, A.E., N.S. Barrett, R.D. Stuart-Smith, N.J. Holbrook, P.A. Thompson, and G.J. Edgar. 2014. Resilience and signatures of tropicalization in protected reef fish communities. Nature Climate Change 4(1):62–67, https://doi.org/10.1038/nclimate2062.
Beger, M., H.S. Grantham, R.L. Pressey, K.A. Wilson, E.L. Peterson, D. Dorfman, P.J. Mumby, R. Lourival, D.R. Brumbaugh, and H.P. Possingham. 2010. Conservation planning for connectivity across marine, freshwater, and terrestrial realms. Biological Conservation 143(3):565–575, https://doi.org/10.1016/j.biocon.2009.11.006.
Bohnsack, J. 1990. The Potential of Marine Fishery Reserves for Reef Fish Management in the US Southern Atlantic. NOAA Technical Memorandum NMFS-SEFC-261, 55 pp, https://doi.org/10.13140/2.1.3007.9369.
Botsford, L.W., A. Hastings, and S.D. Gaines. 2001. Dependence of sustainability on the configuration of marine reserves and larval dispersal distance. Ecology Letters 4:144–150, https://doi.org/10.1046/j.1461-0248.2001.00208.x.
Botsford, L.W., J.W. White, M.H. Carr, and J.E. Caselle. 2014a. Marine protected area networks in California, USA. Pp. 205–251 in Marine Managed Areas and Fisheries. M.L. Johnson, and J. Sandell, eds, Advances in Marine Biology, vol. 69, https://doi.org/10.1016/B978-0-12-800214-8.00006-2.
Botsford, L.W., M.D. Holland, J.C. Field, and A. Hastings. 2014b. Cohort resonance: A significant component of fluctuations in recruitment, egg production, and catch of fished populations. ICES Journal of Marine Science 71(8):2,158–2,170, https://doi.org/10.1093/icesjms/fsu063.
Bruno, J.F., I.M. Côté, and L.T. Toth. 2019. Climate change, coral loss, and the curious case of the parrotfish paradigm: Why don’t marine protected areas improve reef resilience? Annual Review of Marine Science 11(1):307–334, https://doi.org/10.1146/annurev-marine-010318-095300.
Cabral, R.B., S.D. Gaines, M.T. Lim, M.P. Atrigenio, S.S. Mamauag, G.C. Pedemonte, and P.M. Aliño. 2016. Siting marine protected areas based on habitat quality and extent provides the greatest benefit to spatially structured metapopulations. Ecosphere 7(11):e01533, https://doi.org/10.1002/ecs2.1533.
Carr, M.H., S.P. Robinson, C. Wahle, G. Davis, S. Kroll, S. Murray, E.J. Schumacker, and M. Williams. 2017. The central importance of ecological spatial connectivity to effective coastal marine protected areas and to meeting the challenges of climate change in the marine environment. Aquatic Conservation: Marine and Freshwater Ecosystems 27:6–29, https://doi.org/10.1002/aqc.2800.
Carr, M.H., K.L.C. Bell, P. Leary, H.L. Sagar, and S. Tucker. 2018. How MPAs Enhance the Resilience of Coastal Marine Ecosystems and Their Services: A Supplemental Report by the Marine Protected Areas Federal Advisory Committee’s Ecosystem Team. 24 pp.
Caselle, J.E., A. Rassweiler, S.L. Hamilton, and R.R. Warner. 2015. Recovery trajectories of kelp forest animals are rapid yet spatially variable across a network of temperate marine protected areas. Scientific Reports 5:1–14, https://doi.org/10.1038/srep14102.
Caselle, J.E., K. Davis, and L.M. Marks. 2018. Marine management affects the invasion success of a non-native species in a temperate reef system in California, USA. Ecology Letters 21(1):43–53, https://doi.org/10.1111/ele.12869.
Caveen, A.J., T.S. Gray, S.M. Stead, and N.V.C. Polunin. 2013. MPA policy: What lies behind the science? Marine Policy 37(1):3–10, https://doi.org/10.1016/j.marpol.2012.04.005.
Chavez, F.P., C. Costello, D. Aseltine-Neilson, H. Doremus, J.C. Field, S.D. Gaines, M. Hall-Arber, N.J. Mantua, B. McCovey, C. Pomeroy, and others (California Ocean Protection Council Science Advisory Team Working Group). 2017. Readying California Fisheries for Climate Change. California Ocean Science Trust, Oakland, CA, 58 pp.
Christie, M.R., B.N. Tissot, M.A. Albins, J.P. Beets, Y. Jia, D.M. Ortiz, S.E. Thompson, and M.A. Hixon. 2010. Larval connectivity in an effective network of marine protected areas. PLOS ONE 5(12):e15715, https://doi.org/10.1371/journal.pone.0015715.
Cinner, J.E. 2007. Designing marine reserves to reflect local socioeconomic conditions: Lessons from long-enduring customary management systems. Coral Reefs 26(4):1,035–1,045, https://doi.org/10.1007/s00338-007-0213-2.
Cinner, J.E., E. Maire, C. Huchery, M.A. MacNeil, N.A.J. Graham, C. Mora, T.R. McClanahan, M.L. Barnes, J.N. Kittinger, C.C. Hicks, and others. 2018. Gravity of human impacts mediates coral reef conservation gains. Proceedings of the National Academy of Sciences of the United States of America 115(27):E6116–E6125, https://doi.org/10.1073/pnas.1708001115.
Claisse, J.T., C.A. Blanchette, J.E. Dugan, J.P. Williams, J. Freiwald, D.J. Pondella, I.I. Nicholas, K.S. David, K. Davis, L.A. Zahn, and others. 2018. Biogeographic patterns of communities across diverse marine ecosystems in southern California. Marine Ecology 39:e12453, https://doi.org/10.1111/maec.12453.
Claudet, J., ed. 2011. Marine Protected Areas: A Multidisciplinary Approach. Cambridge University Press, Cambridge, UK, 373 pp.
Cohen, P.J., and S.J. Foale. 2013. Sustaining small-scale fisheries with periodically harvested marine reserves. Marine Policy 37(1):278–287, https://doi.org/10.1016/j.marpol.2012.05.010.
Costello, C., A. Rassweiler, D. Siegel, G. De Leo, F. Micheli, and A. Rosenberg. 2010. The value of spatial information in MPA network design. Proceedings of the National Academy of Sciences of the United States of America 107(43):18,294–18,299, https://doi.org/10.1073/pnas.0908057107.
Dugan, J.E., and G.E. Davis. 1993. Introduction to the international symposium on marine harvest refugia. Canadian Journal of Fisheries and Aquatic Sciences 50(9):1,991–1,992, https://doi.org/10.1139/f93-222.
Edgar, G.J., R.D. Stuart-Smith, T.J. Willis, S. Kininmonth, S.C. Baker, S. Banks, N.S. Barrett, M.A. Becerro, A.T.F. Bernard, J. Berkhout, and others. 2014. Global conservation outcomes depend on marine protected areas with five key features. Nature 506(7487):216–220, https://doi.org/10.1038/nature13022.
Field, J.C., A.E. Punt, R.D. Methot, and C.J. Thomson. 2006. Does MPA mean ‘Major Problem for Assessments’? Considering the consequences of place-based management systems. Fish and Fisheries 7:284–302, https://doi.org/10.1111/j.1467-2979.2006.00226.x.
Fodrie, F.J., and L.A. Levin. 2008. Linking juvenile habitat utilization to population dynamics of California halibut. Limnology and Oceanography 53(2):799–812, https://doi.org/10.4319/lo.2008.53.2.0799.
Fox, E., M. Miller-Henson, J. Ugoretz, M. Weber, M. Gleason, J. Kirlin, M. Caldwell, and S. Mastrup. 2013. Enabling conditions to support marine protected area network planning: California’s Marine Life Protection Act Initiative as a case study. Ocean & Coastal Management 74:14–23, https://doi.org/10.1016/j.ocecoaman.2012.07.005.
Fox, A.D., L. Henry, D.W. Corne, and J.M. Roberts. 2016. Sensitivity of marine protected area network connectivity to atmospheric variability. Royal Society Open Science 3:160494, https://doi.org/10.1098/rsos.160494.
Freiwald, J. 2012. Movement of adult temperate reef fishes off the west coast of North America. Canadian Journal of Fisheries and Aquatic Sciences 69(8):1,362–1,374, https://doi.org/10.1139/f2012-068.
Friedlander, A.M., J.M. Shackeroff, and J.N. Kittinger. 2013. Customary marine resource knowledge and use in contemporary Hawai‘i. Pacific Science 67(3):441–460, https://doi.org/10.2984/67.3.10.
Gaines, S.D., B. Gaylord, and J.L. Largier. 2003. Avoiding current oversights in marine reserve design. Ecological Applications 13(sp1):32–46, https://doi.org/10.1890/1051-0761(2003)013[0032:ACOIMR]2.0.CO;2.
Gaines, S.D., C. White, M.H. Carr, and S.R. Palumbi. 2010. Designing marine reserve networks for both conservation and fisheries management. Proceedings of the National Academy of Sciences of the United States of America 107(43):18,286–18,293, https://doi.org/10.1073/pnas.0906473107.
Game, E.T., M. Bode, E. McDonald-Madden, H.S. Grantham, and H.P. Possingham. 2009. Dynamic marine protected areas can improve the resilience of coral reef systems. Ecology Letters 12(12):1,336–1,345, https://doi.org/10.1111/j.1461-0248.2009.01384.x.
Gleason, M., J. Kirlin, and E. Fox. 2013. California’s marine protected area network planning process: Introduction to the special issue. Ocean & Coastal Management 74:1–2, https://doi.org/10.1016/j.ocecoaman.2012.08.022.
Goetze, J.S., J. Claudet, F. Januchowski-Hartley, T.J. Langlois, S.K. Wilson, C. White, R. Weeks, and S.D. Jupiter. 2018. Demonstrating multiple benefits from periodically harvested fisheries closures. Journal of Applied Ecology 55:1,102–1,113, https://doi.org/10.1111/1365-2664.13047.
Green, A.L., L. Fernandes, G. Almany, R. Abesamis, E. McLeod, P.M. Aliño, A.T. White, R. Salm, J. Tanzer, and R.L. Pressey. 2014. Designing marine reserves for fisheries management, biodiversity conservation, and climate change adaptation. Coastal Management 42(2):143–159, https://doi.org/10.1080/08920753.2014.877763.
Grorud-Colvert, K., J. Claudet, B.N. Tissot, J.E. Caselle, M.H. Carr, J.C. Day, A.M. Friedlander, S.E. Lester, T. Lison de Loma, D. Malone, and others. 2014. Marine protected area networks: Assessing whether the whole is greater than the sum of its parts. PLOS ONE 9(8):e102298, https://doi.org/10.1371/journal.pone.0102298.
Grüss, A., J. Robinson, S.S. Heppell, S.A. Hellell, and B.X. Semmens. 2014. Conservation and fisheries effects of spawning aggregation marine protected areas: What we know, where we should go, and what we need to get there. ICES Journal of Marine Science 71(7):1,515–1,534, https://doi.org/10.1093/icesjms/fsu038.
Halpern, B.S., S.E. Lester, and K.L. McLeod. 2010a. Placing marine protected areas onto the ecosystem-based management seascape. Proceedings of the National Academy of Sciences of the United States of America 107(43):18,312–18,317, https://doi.org/10.1073/pnas.0908503107.
Halpern, B.S., S.E. Lester, and J.B. Kellner. 2010b. Spillover from marine reserves and the replenishment of fished stocks. Environmental Conservation 36(4):268–276, https://doi.org/10.1017/S0376892910000032.
Hamilton, S.L., J.E. Caselle, D.P. Malone, and M.H. Carr. 2010. Incorporating biogeography into evaluations of the Channel Islands marine reserve network. Proceedings of the National Academy of Sciences of the United States of America 107(43):1,822–1,827, https://doi.org/10.1073/pnas.0908091107.
Harrison, H.B., D.H. Williamson, R.D. Evans, G.R. Almany, S.R. Thorrold, G.R. Russ, K.A. Feldheim, L. van Herwerden, S. Planes, M. Srinivasan, and others. 2012. Larval export from marine reserves and the recruitment benefit for fish and fisheries. Current Biology 22(11):1,023–1,028, https://doi.org/10.1016/j.cub.2012.04.008.
Hart, D.R. 2003. Yield- and biomass-per-recruit analysis for rotational fisheries, with an application to the Atlantic sea scallop (Placopecten magellanicus). Fishery Bulletin 101(1):44–57.
Hastings, A., and L.W. Botsford. 1999. Equivalence in yield from marine reserves and traditional fisheries management. Science 284:1,537–1,538, https://doi.org/10.1126/science.284.5419.1537.
Hopf, J.K., G.P. Jones, D.H. Williamson, and S.R. Connolly. 2016. Fishery consequences of marine reserves: Short-term pain for longer-term gain. Ecological Applications 26(3):818–829, https://doi.org/10.1890/15-0348.
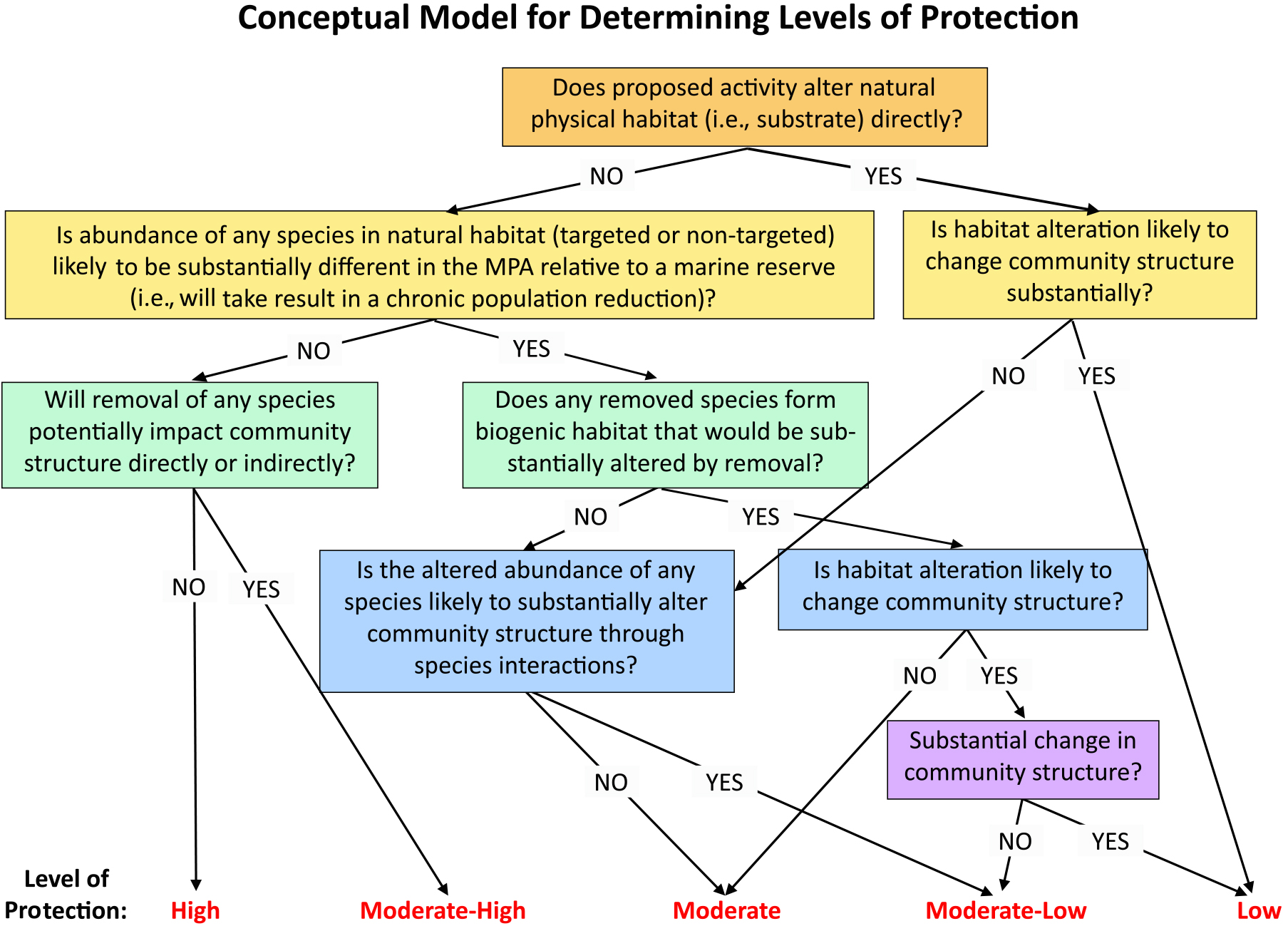
IUCN (International Union for Conservation of Nature). 2018. Applying IUCN’s Global Conservation Standards to Marine Protected Areas (MPAs). Gland, Switzerland, 4 pp.
Johannes, R.E. 1978. Traditional marine conservation methods in Oceania and their demise. Annual Review of Ecology and Systematics 9:349–364, https://doi.org/10.1146/annurev.es.09.110178.002025.
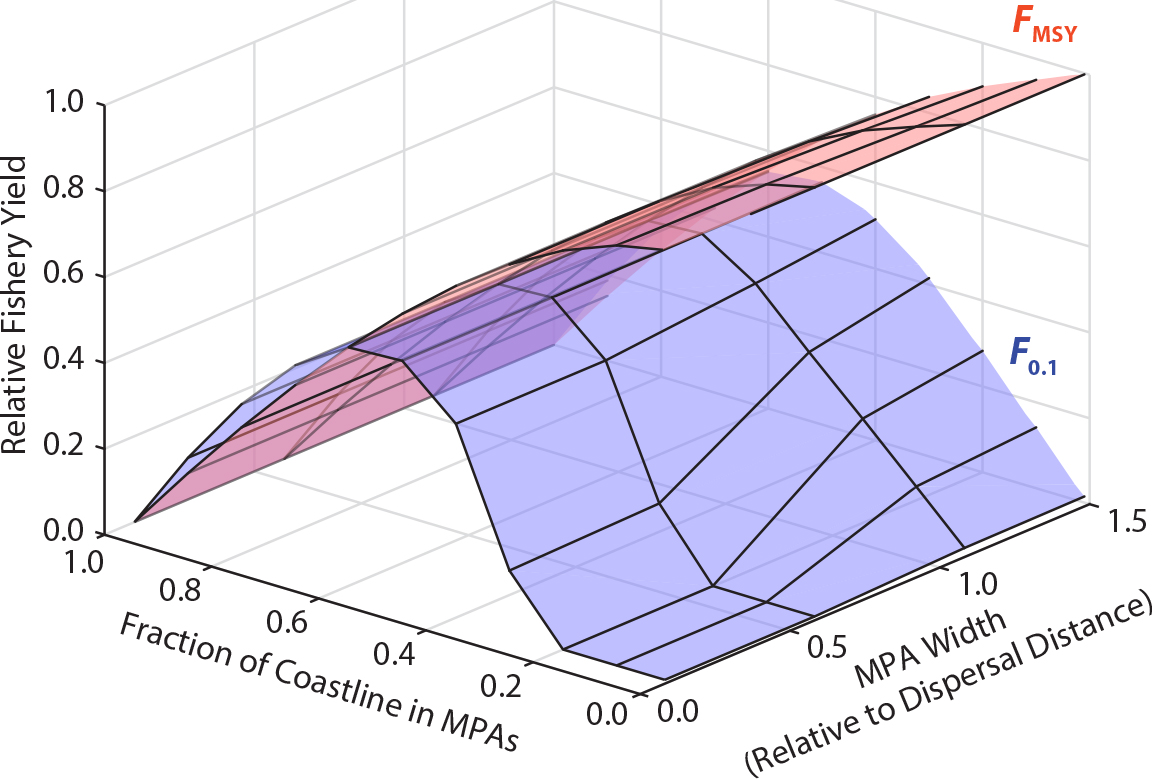
Kaplan, D.M., and L.W. Botsford. 2005. Effects of variability in spacing of coastal marine reserves on fisheries yield and sustainability. Canadian Journal of Fisheries and Aquatic Sciences 62:905–912, https://doi.org/10.1139/f04-243.
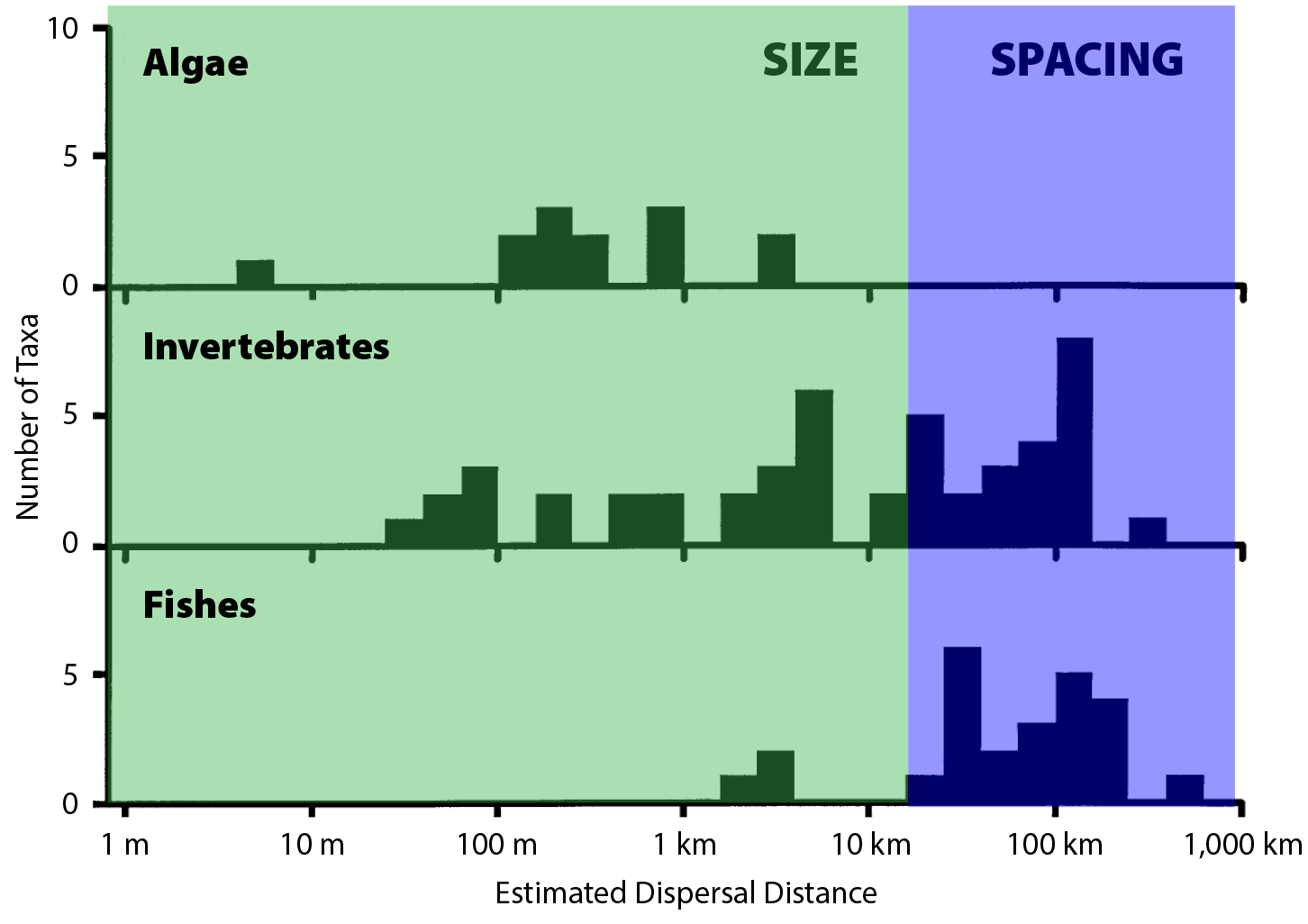
Kinlan, B.P., and S.D. Gaines. 2003. Propagule dispersal in marine and terrestrial environments: A community perspective. Ecology 84(8):2,007–2,020, https://doi.org/10.1890/01-0622.
Kinlan, B.P., S.D. Gaines, and S.E. Lester. 2005. Propagule dispersal and the scales of marine community process. Diversity and Distributions 11(2):139–148, https://doi.org/10.1111/j.1366-9516.2005.00158.x.
Largier, J.L. 2003. Considerations in estimating larval dispersal distance from oceanographic data. Ecological Applications 13(sp1):71–89, https://doi.org/10.1890/1051-0761(2003)013[0071:CIELDD]2.0.CO;2.
Leslie, H.M. 2005. A synthesis of marine conservation planning approaches. Conservation Biology 19(6):1,701–1,713, https://doi.org/10.1111/j.1523-1739.2005.00268.x.
Lester, S.S.E., B.B.S. Halpern, K. Grorud-Colvert, J. Lubchenco, B.I.B. Ruttenberg, S.D.S. Gaines, S. Airamé, and R.R.R. Warner. 2009. Biological effects within no-take marine reserves: A global synthesis. Marine Ecology Progress Series 384:33–46, https://doi.org/10.3354/meps08029.
Ling, S.D., and C.R. Johnson. 2012. Marine reserves reduce risk of climate-driven phase shift by reinstating size-and habitat-specific trophic interactions. Ecological Applications 22(4):1,232–1,245, https://doi.org/10.1890/11-1587.1.
Lubchenco, J., S.R. Palumbi, S.D. Gaines, and S. Andelman. 2003. Plugging a hole in the ocean: The emerging science of marine reserves. Ecological Applications 13(sp1):3–7, https://doi.org/10.1890/1051-0761(2003)013[0003:PAHITO]2.0.CO;2.
McGilliard, C.R., R. Hilborn, A. MacCall, A.E. Punt, and J.C. Field. 2011. Can we use information from marine protected areas to inform management of small-scale, data-poor stocks? ICES Journal of Marine Science 68(1):201–211, https://doi.org/10.1093/icesjms/fsq151.
McLeod, E., R. Salm, A. Green, and J. Almany. 2009. Designing marine protected area networks to address the impacts of climate change. Frontiers in Ecology and the Environment 7(7):362–370, https://doi.org/10.1890/070211.
Moffitt, E.A., J. W. White, and L.W. Botsford. 2011. The utility and limitations of size and spacing guidelines for designing marine protected area (MPA) networks. Biological Conservation 144(1):306–318, https://doi.org/10.1016/j.biocon.2010.09.008.
Nagelkerken, I., M.G.G. Grol, and P.J. Mumby. 2012. Effects of marine reserves versus nursery habitat availability on structure of reef fish communities. PLOS ONE 7(6):e36906, https://doi.org/10.1371/journal.pone.0036906.
Nickols, K.J., J.W. White, D.P. Malone, M.H. Carr, R.M. Starr, M.L. Baskett, A. Hastings, and L.W. Botsford. In press. Setting ecological expectations for adaptive management of marine protected areas. Journal of Applied Ecology, https://doi.org/10.1111/1365-2664.13463.
Palumbi, S.R. 2003. Population genetics, demographic connectivity and the design of marine reserves. Ecological Applications 13(sp1):146–158, https://doi.org/10.1890/1051-0761(2003)013[0146:PGDCAT]2.0.CO;2.
Rassweiler, A., C. Costello, and D.A. Siegel. 2012. Marine protected areas and the value of spatially optimized fishery management. Proceedings of the National Academy of Sciences of the United States of America 109(29):11,884–11,889, https://doi.org/10.1073/pnas.1116193109.
Roberts, C.M., J.A. Bohnsack, F. Gell, J.P. Hawkins, and R. Goodridge. 2001. Effects of marine reserves on adjacent fisheries. Science 294(5548):1,920–1,923, https://doi.org/10.1126/science.294.5548.1920.
Saarman, E.T., M. Gleason, J. Ugoretz, S. Airamé, M.H. Carr, E. Fox, A. Frimodig, T. Mason, and J. Vasques. 2013. The role of science in supporting marine protected area network planning and design in California. Ocean & Coastal Management 74:45–56, https://doi.org/10.1016/j.ocecoaman.2012.08.021.
Tupper, M.H. 2007. Spillover of commercially valuable reef fishes from marine protected areas in Guam, Micronesia. Fishery Bulletin 105(5):527–537, https://www.worldfishcenter.org/content/spillover-commercially-valuable-reef-fishes-marine-protected-areas-guam-micronesia.
Watson, J.R., S. Mitarai, D.A. Siegel, J.E. Caselle, C. Dong, and J.C. McWilliams. 2010. Realized and potential larval connectivity in the southern California bight. Marine Ecology Progress Series 401:31–48, https://doi.org/10.3354/meps08376.
Weible, C., P.A. Sabatier, and M. Lubell. 2004. A comparison of a collaborative and top-down approach to the use of science in policy: Establishing marine protected areas in California. Policy Studies Journal 32(2):187–208, https://doi.org/10.1111/j.1541-0072.2004.00060.x.
White, C., and B.E. Kendall. 2007. A reassessment of equivalence in yield from marine reserves and traditional fisheries management. Oikos 116:2,039–2,043, https://doi.org/10.1111/j.2007.0030-1299.16167.x.
White, J.W., L.W. Botsford, E.A. Moffitt, and D.T. Fischer. 2010a. Decision analysis for designing marine protected areas for multiple species with uncertain fishery status. Ecological Applications 20(6):1,523–1,541, https://doi.org/10.1890/09-0962.1.
White, J.W., L.W. Botsford, A. Hastings, and J.L. Largier. 2010b. Population persistence in marine reserve networks: Incorporating spatial heterogeneities in larval dispersal. Marine Ecology Progress Series 398:49–67, https://doi.org/10.3354/meps08327.
White, J.W., L.W. Botsford, M.L. Baskett, L.A.K. Barnett, R.J. Barr, and A. Hastings. 2011. Linking models with monitoring data for assessing performance of no-take marine reserves. Frontiers in Ecology and the Environment 9(7):390–399, https://doi.org/10.1890/100138.
White, J.W., A.J. Scholz, A. Rassweiler, C. Steinback, L.W. Botsford, S. Kruse, C. Costello, S. Mitarai, D.A. Siegel, P.T. Drake, and others. 2013a. A comparison of approaches used for economic analysis in marine protected area network planning in California. Ocean & Coastal Management 74:77–89, https://doi.org/10.1016/j.ocecoaman.2012.06.006.
White, J.W., L.W. Botsford, A. Hastings, M.L. Baskett, D.M. Kaplan, and L.A.K. Barnett. 2013b. Transient responses of fished populations to marine reserve establishment. Conservation Letters 6(3):180–191, https://doi.org/10.1111/j.1755-263X.2012.00295.x.
White, J.W., M.H. Carr, J.E. Caselle, L. Washburn, C.B. Woodson, S.R. Palumbi, P.M. Carlson, R.R. Warner, B.A. Menge, J.A. Barth, and others. 2019. Connectivity, dispersal, and recruitment: Connecting benthic communities and the coastal ocean. Oceanography 32(3):50–59, https://doi.org/10.5670/oceanog.2019.310.
Young, M.A., L.W. Wedding, and M.H. Carr. 2017. Applying landscape ecology to evaluate the design of marine protected area networks. Pp. 429–452 in Seascape Ecology: Taking Landscape Ecology into the Sea. S. Pittman, ed., John Wiley & Sons, Chichester, UK.