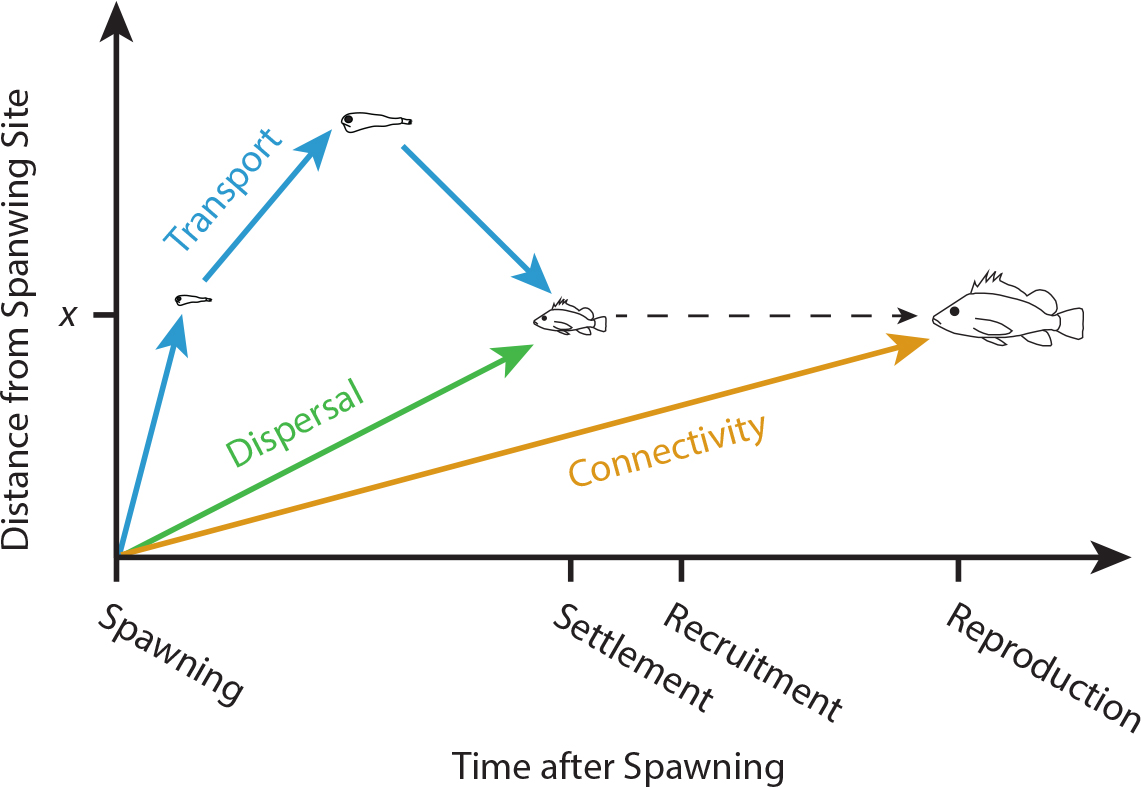
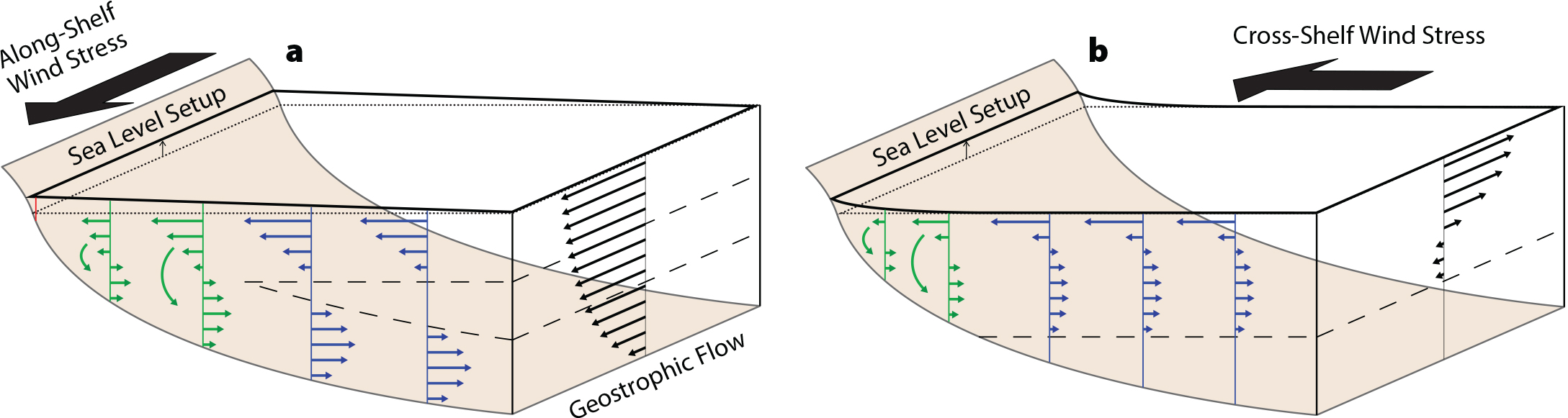
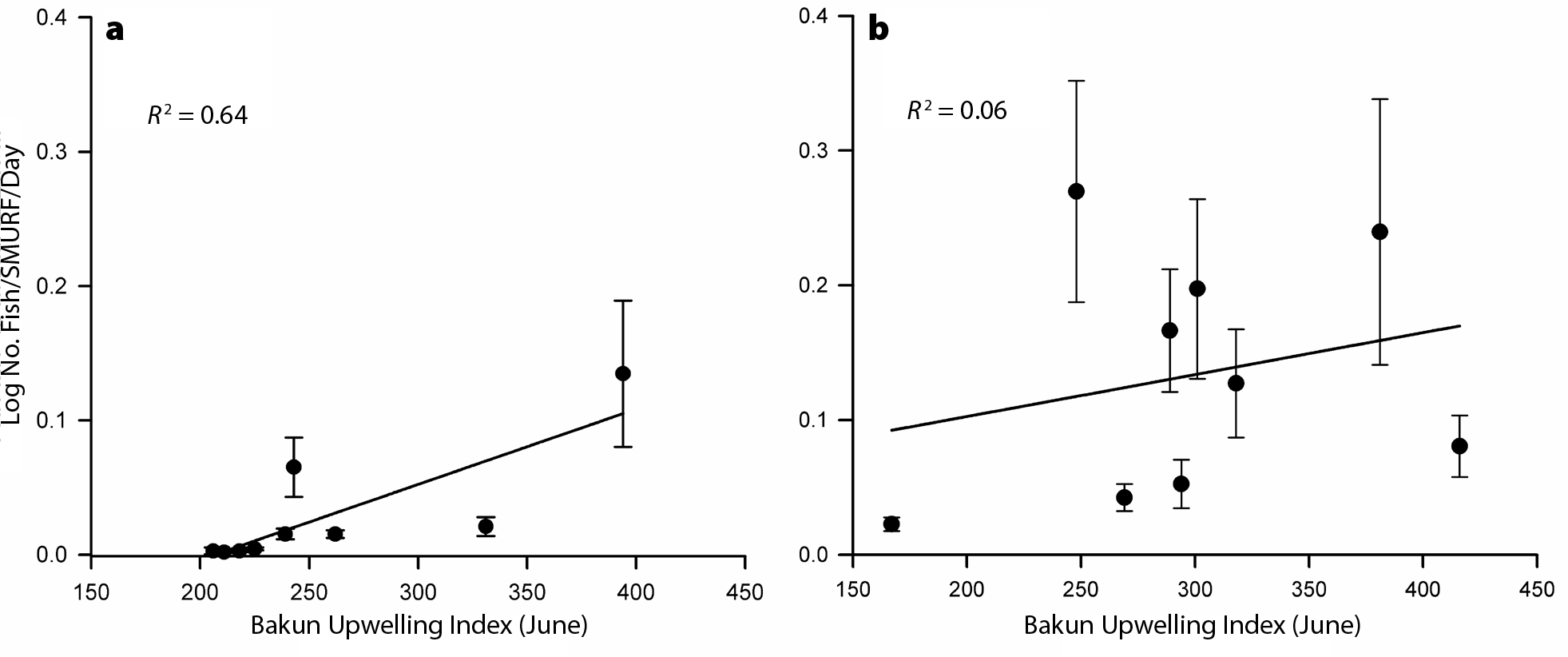
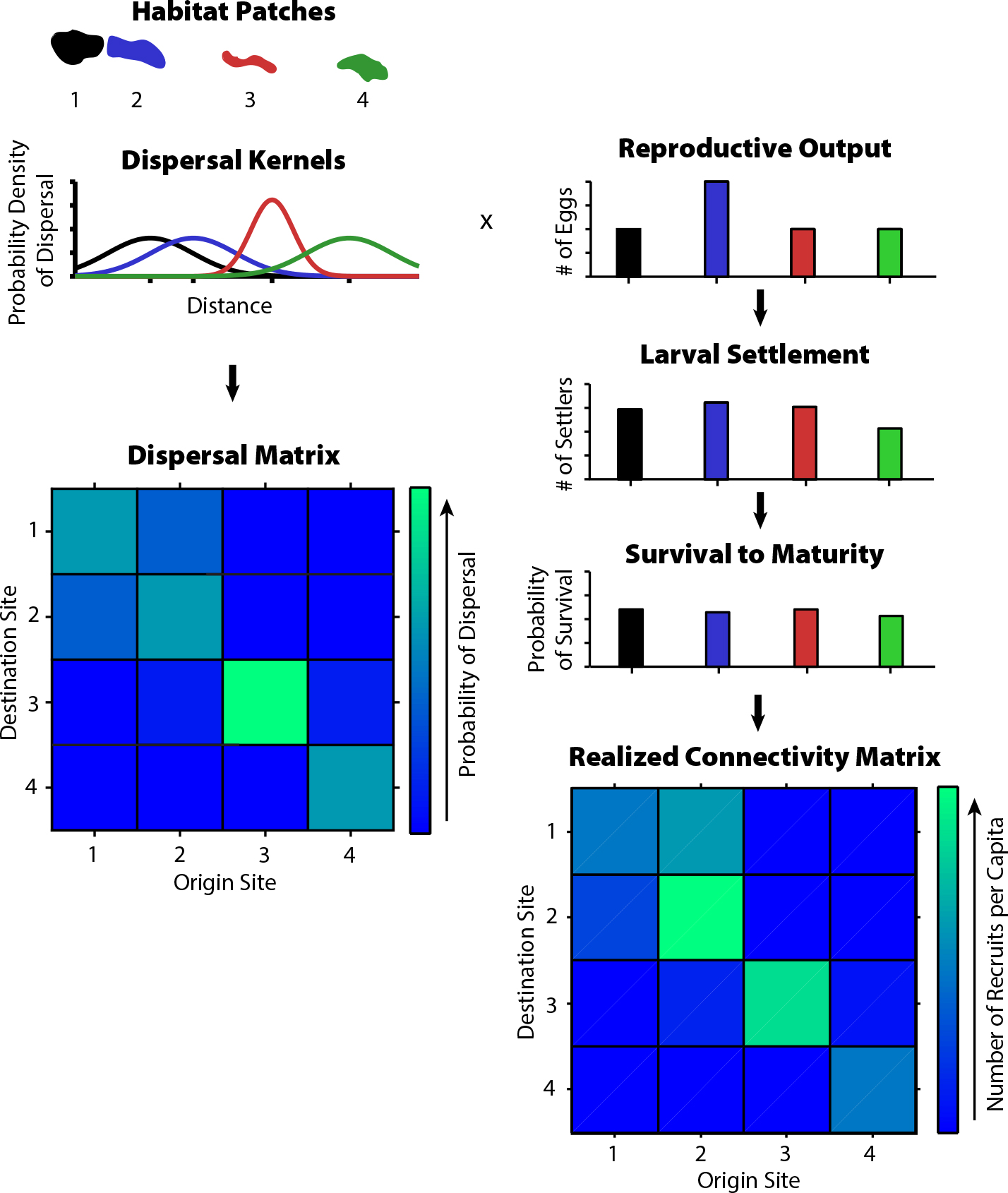
Austin, J.A., and S.J. Lentz. 2002. The inner shelf response to wind-driven upwelling and downwelling. Journal of Physical Oceanography 32:2,171–2,193, https://doi.org/10.1175/1520-0485(2002)032<2171:TISRTW>2.0.CO;2.
Baetscher, D.S., E.C. Anderson, E.A. Gilbert-Horvath, D. Malone, E.T. Saarman, M.H. Carr, and J.C. Garza. 2019. Dispersal of a nearshore marine fish connects marine reserves and adjacent fished areas along an open coast. Molecular Ecology 28:1–13, https://doi.org/10.1111/mec.15044.
Barneche, D.R., D.R. Robertson, C.R. White, and D.J. Marshall. 2018. Fish reproductive-energy output increases disproportionately with body size. Science 360:642–645, https://doi.org/10.1126/science.aao6868.
Browman, H.I. 2014. Commemorating 100 years since Hjort’s 1914 treatise on fluctuations in the great fisheries of northern Europe: Where we have been, where we are, and where we are going. ICES Journal of Marine Science 71:1,989–1,992, https://doi.org/10.1093/icesjms/fsu159.
Burgess, S.C., K.J. Nickols, C.D. Griesemer, L.A.K. Barnett, A.G. Dedrick, E.V. Satterthwaite, L. Yamane, S.G. Morgan, J.W. White, and L.W. Botsford. 2014. Beyond connectivity: How empirical methods can quantify population persistence to improve marine protected area design. Ecological Applications 24:257–270, https://doi.org/10.1890/13-0710.1.
Carr, M.H., J.W. White, E. Saarman, J. Lubchenco, K. Milligan, and J.E. Caselle. 2019. Marine protected areas exemplify the evolution of science and policy. Oceanography 32(3):94–103, https://doi.org/10.5670/oceanog.2019.315.
Caselle, J.E., B.P. Kinlan, and R.R. Warner. 2010a. Temporal and spatial scales of influence on nearshore fish settlement in the Southern California Bight. Bulletin of Marine Science 86:355–385.
Caselle, J.E., J.R. Wilson, M.H. Carr, D.P. Malone, and D.E. Wendt. 2010b. Can we predict interannual and regional variation in delivery of pelagic juveniles to nearshore populations of rockfishes (genus Sebastes) using simple proxies of ocean conditions? CalCOFI Reports 51:1–16.
Cushing, D.H. 1990. Plankton production and year-class strength in fish populations: An update of the match/mismatch hypothesis. Advances in Marine Biology 26:249–293, https://doi.org/10.1016/S0065-2881(08)60202-3.
Cowen, R.K., and S. Sponaugle. 2009. Larval dispersal and marine population connectivity. Annual Review of Marine Science 1:443–466,
https://doi.org/10.1146/annurev.marine.010908.163757.
De Wit, P., L. Rogers-Bennett, R.M. Kudela, and S.R. Palumbi. 2014. Forensic genomics as a novel tool for identifying the causes of mass mortality events. Nature Communications 5:3652, https://doi.org/10.1038/ncomms4652.
Dingeldein, A., and J.W. White. 2016. Larval traits carry over to affect post-settlement behaviour in a common coral reef fish. Journal of Animal Ecology 85:903–914, https://doi.org/10.1111/1365-2656.12506.
Drake, P.T., C.A. Edwards, S.G. Morgan, and E.P. Dever. 2013. Influence of larval behavior on transport and population connectivity in a realistic simulation of the California Current System. Journal of Marine Research 71:317–350, https://doi.org/10.1357/002224013808877099.
Durant, J.M., D.Ø. Hjermann, T. Falkenhaug, D. Gifford, L.J. Naustvoll, B. Sullivan, G. Beaugrand, and N.C. Stenseth. 2013. Extension of the match-mismatch hypothesis to predator-controlled systems. Marine Ecology Progress Series 474:43–52, https://doi.org/10.3354/meps10089.
Figueira, W.F., and L.B. Crowder. 2006. Defining patch contribution in source-sink metapopulations: The importance of including dispersal and its relevance to marine systems. Population Ecology 48:215–224, https://doi.org/10.1007/s10144-006-0265-0.
Fouzai, N., A.F. Opdal, C. Jørgensen, and Ø. Fiksen. 2015. Effects of temperature and food availability on larval cod survival: A model for behaviour in vertical gradients. Marine Ecology Progress Series 529:199–212, https://doi.org/10.3354/meps11326.
Garavelli, L., J.W. White, I. Chollet, S.J. Box, and L.M. Chérubin. 2018. Population models reveal unexpected patterns of local persistence despite widespread larval dispersal in a highly exploited species. Conservation Letters 11(5):e12567, https://doi.org/10.1111/conl.12567.
Gerber, L.R., M. Del Mar Mancha-Cisneros, M.I. O’Connor, and E.R. Selig. 2014. Climate change impacts on connectivity in the ocean: Implications for conservation. Ecosphere 5:33, https://doi.org/10.1890/ES13-00336.1.
Gruenthal, K.M., L.K. Acheson, and R.S. Burton. 2007. Genetic structure of natural populations of California red abalone (Haliotis rufescens) using multiple genetic markers. Marine Biology 152:1,237–1,248, https://doi.org/10.1007/s00227-007-0771-4.
Hamilton, S.L., J. Regetz, and R.R. Warner. 2008. Postsettlement survival linked to larval life in a marine fish. Proceedings of the National Academy of Sciences of the United States of America 105:1,561–1,566, https://doi.org/10.1073/pnas.0707676105.
Hare, J.A., and R.K. Cowen. 1997. Size, growth, development, and survival of the planktonic larvae of Pomatomus saltatrix (Pisces: Pomatomidae). Ecology 78:2,415–2,431, https://doi.org/10.1890/0012-9658(1997)078[2415:SGDASO]2.0.CO;2.
He, X., J.C. Field, S.G. Beyer, and S.M. Sogard. 2015. Effects of size-dependent relative fecundity specifications in fishery stock assessments. Fisheries Research 165:54–62, https://doi.org/10.1016/j.fishres.2014.12.023.
Huyer, A. 1983. Coastal upwelling in the California Current system. Progress in Oceanography 12:259–284, https://doi.org/10.1016/0079-6611(83)90010-1.
Johnson, D.W., M.R. Christie, T.J. Pusack, C.D. Stallings, and M.A. Hixon. 2018. Integrating larval connectivity with local demography reveals regional dynamics of a marine metapopulation. Ecology 99:1,419–1,429, https://doi.org/10.1002/ecy.2343.
Kallimanis, A.S., W.E. Kunin, J.M. Halley, and S.P. Sgardelis. 2005. Metapopulation extinction risk under spatially autocorrelated disturbance. Conservation Biology 19:534–546, https://doi.org/10.1111/j.1523-1739.2005.00418.x.
Kim, S.Y., B.D. Cornuelle, E.J. Terrill, B. Jones, L. Washburn, M.A. Moline, J.D. Paduan, N. Garfield, J.L. Largier, G. Crawford, and P.M. Kosro. 2013. Poleward propagating subinertial alongshore surface currents off the US west coast. Journal of Geophysical Research 118:6,791–6,806, https://doi.org/10.1002/jgrc.20400.
Leis, J.M. 2007. Behaviour as input for modelling dispersal of fish larvae: Behaviour, biogeography, hydrodynamics, ontogeny, physiology and phylogeny meet hydrography. Marine Ecology Progress Series 347:185–193, https://doi.org/10.3354/meps06977.
Leising, A.W., I.D. Schroder, S.J. Bograd, J. Abell, R. Durazo, G. Gaxiola-Castro, E.P. Bjorkstedt, F. Field, K. Sakuma, R.R. Robertson, and others. 2015. State of the California Current 2014–15: Impacts of the warm-water ‘Blob.’ CalCOFI Reports 56:31–68.
Lentz, S J., and M.R. Fewings. 2012. The wind- and wave-driven inner-shelf circulation. Annual Review of Marine Science 4:317–343, https://doi.org/10.1146/annurev-marine-120709-142745.
Menge, B.A., T.C. Gouhier, T. Freidenburg, and J. Lubchenco. 2011. Linking long-term, large-scale climatic and environmental variability to patterns of marine invertebrate recruitment: Toward explaining “unexplained” variation. Journal of Experimental Marine Biology and Ecology 400:236–249, https://doi.org/10.1016/j.jembe.2011.02.003.
Menge, B.A., K. Milligan, J.E. Caselle, J.A. Barth, C.A. Blanchette, M.H. Carr, F. Chan, R.K. Cowen, M. Denny, S.D. Gaines, and others. 2019. PISCO: Advances made through the formation of a large-scale, long-term consortium for integrated understanding of coastal ecosystem dynamics. Oceanography 32(3):16–25, https://doi.org/10.5670/oceanog.2019.307.
Morgan, S.G. 2014. Behaviorally mediated larval transport in upwelling systems. Advances in Oceanography 2014:364214, https://doi.org/10.1155/2014/364214.
Nickols, K.J., B. Gaylord, and J.L. Largier. 2012. The coastal boundary layer: Predictable current structure decreases alongshore transport and alters scales of dispersal. Marine Ecology Progress Series 464:17–35, https://doi.org/10.3354/meps09875.
Nickols, K.J., J.W. White, J.L. Largier, and B. Gaylord. 2015. Marine population connectivity: Reconciling large-scale dispersal and high self-retention. The American Naturalist 185:196–211, https://doi.org/10.1086/679503.
Parrish, R.H., C.S. Nelson, and A. Bakun. 1981. Transport mechanisms and reproductive success of fishes in the California Current. Biological Oceanography 1:175–203.
Pineda, J., J.A. Hare, and S. Sponaugle. 2007. Larval transport and dispersal in the coastal ocean: Consequences for population connectivity. Oceanography 20:22–39, https://doi.org/10.5670/oceanog.2007.27.
Shanks, A.L., and G.L. Eckert. 2005. Population persistence of California Current fishes and benthic crustaceans: A marine drift paradox. Ecological Monographs 75:505–524, https://doi.org/10.1890/05-0309.
Shanks, A.L., and G.C. Roegner. 2007. Recruitment limitation in Dungeness crab populations is driven by variation in atmospheric forcing. Ecology 88:1,726–1,737, https://doi.org/10.1890/06-1003.1.
Shanks, A.L., S.G. Morgan, J. MacMahan, and A.J.H.M. Reniers. 2010. Surf zone physical and morphological regime as determinants of temporal and spatial variation in larval recruitment. Journal of Experimental Marine Biology and Ecology 392:140–150, https://doi.org/10.1016/j.jembe.2010.04.018.
Shima, J.S., and S.E. Swearer. 2010. The legacy of dispersal: Larval experience shapes persistence later in the life of a reef fish. Journal of Animal Ecology 79:1,308–1,314, https://doi.org/10.1111/j.1365-2656.2010.01733.x.
Siegel, D.A., S. Mitarai, C.J. Costello, S.D. Gaines, B.E. Kendall, R.R. Warner, and K.B. Winters. 2008. The stochastic nature of larval connectivity among nearshore marine populations. Proceedings of the National Academy of Sciences of the United States of America 105:8,974–8,979, https://doi.org/10.1073/pnas.0802544105.
Sponaugle, S., C. Paris, K.D. Walter, V. Kourafalou, and E. D’Alessandro. 2012. Observed and modeled larval settlement of a reef fish to the Florida Keys. Marine Ecology Progress Series 453:201–212, https://doi.org/10.3354/meps09641.
Staaterman, E., and C.B. Paris. 2013. Modeling larval fish navigation: The way forward. ICES Journal of Marine Science 71:918–924, https://doi.org/10.1093/icesjms/fst103.
Swearer, S.E., J.S. Shima, M.E. Hellberg, S.R. Thorrold, G.P. Jones, D.R. Robertson, S.G. Morgan, K.A. Selkoe, G.M. Ruiz, and R.R. Warner. 2002. Evidence of self-recruitment in demersal marine populations. Bulletin of Marine Science 70:251–271.
Warner, J.C., B. Armstrong, R. He, and J.B. Zambon. 2010. Development of a Coupled Ocean-Atmosphere-Wave-Sediment Transport (COAWST) Modeling System. Ocean Modelling 35:230–244, https://doi.org/10.1016/j.ocemod.2010.07.010.
Warner, R.R., and P.L. Chesson. 1985. Coexistence mediated by recruitment fluctuations: A field guide to the storage effect. The American Naturalist 125:769–787, https://doi.org/10.1086/284379.
Washburn, L., M.R. Fewings, C. Melton, and C. Gotschalk. 2011. The propagating response of coastal circulation due to wind relaxations along the central California coast. Journal of Geophysical Research: Oceans 116:C12028, https://doi.org/10.1029/2011JC007502.
Watson, J.R., D.A. Siegel, B.E. Kendall, S. Mitarai, A. Rassweiler, and S.D. Gaines. 2011. Identifying critical regions in small-world metapopulations. Proceedings of the National Academy of Sciences of the United States of America 108:E907–E913, https://doi.org/10.1073/pnas.1111461108.
Wheeler, S.G., T.W. Anderson, T.W. Bell, S.G. Morgan, and J.A. Hobbs. 2017. Regional productivity predicts individual growth and recruitment of rockfishes in a northern California upwelling system. Limnology and Oceanography 62:754–767, https://doi.org/10.1002/lno.10458.
White, J.W., J.F. Samhouri, A.C. Stier, C.L. Wormald, S.L. Hamilton, and S.A. Sandin. 2010. Synthesizing mechanisms of density dependence in reef fishes: Behavior, habitat configuration, and observational scale. Ecology 91:1,949–1,961, https://doi.org/10.1890/09-0298.1.
White, J.W., and J.F. Samhouri. 2011. Oceanographic coupling across multiple trophic levels shapes source-sink dynamics in marine metacommunities. Oikos 120:1,151–1,164, https://doi.org/10.1111/j.1600-0706.2010.19226.x.
White, J.W., L.W. Botsford, M.L. Baskett, L.A.K. Barnett, R.J. Barr, and A. Hastings. 2011. Linking models and monitoring data for assessing performance of no-take marine reserves. Frontiers in Ecology and the Environment 9:390–399, https://doi.org/10.1890/100138.
White, J.W., M.H. Carr, J.E. Caselle, S.R. Palumbi, R.R. Warner, B.A. Menge, and K. Milligan. 2019. Empirical approaches to measure connectivity. Oceanography 32(3):60–61, https://doi.org/10.5670/oceanog.2019.311.
Woodson, C.B., M.A. McManus, J.A. Tyburczy, J.A. Barth, L. Washburn, J.E. Caselle, M.H. Carr, D.P. Malone, P.T. Raimondi, B.A. Menge, and S.R. Palumbi. 2012. Coastal fronts set recruitment and connectivity patterns across multiple taxa. Limnology and Oceanography 57:582–596, https://doi.org/10.4319/lo.2012.57.2.0582.