Able, K.W., P.C. López-Duarte, F.J. Fodrie, O.P. Jensen, C.W. Martin, B.J. Roberts, J. Valenti, K. O’Connor, and S.C. Halbert. 2015. Fish assemblages in Louisiana salt marshes: Effects of the Macondo oil spill. Estuaries and Coasts 38:1,385–1,398, https://doi.org/10.1007/s12237-014-9890-6.
Almeda, R., S. Baca, C. Hyatt, and E.J. Buskey. 2014a. Ingestion and sublethal effects of physically and chemically dispersed crude oil on marine planktonic copepods. Ecotoxicology 23:988–1,003, https://doi.org/10.1007/s10646-014-1242-6.
Alameda, R., S. Bona, C. Foster, and E.J. Buskey. 2014b. Dispersant Corexit 9500A and chemically dispersed crude oil decreases the growth rates of meroplanktonic barnacle nauplii (Amphibalanus improvisus) and tornaria larvae (Schizocardium sp.). Marine Environmental Research 99:212e217, https://doi.org/10.1016/j.marenvres.2014.06.007.
Almeda, R., T.E. Harvey, T.L. Connelly, S. Baca, and E.J. Buskey. 2016. Influence of UVB radiation on the lethal and sublethal toxicity of dispersed crude oil to planktonic copepod nauplii. Chemosphere 152:446–458, https://doi.org/10.1016/j.chemosphere.2016.02.129.
Almeda, R., S. Cosgrove, and E.J. Buskey. 2018. Oil spills and dispersants can cause the initiation of potentially harmful dinoflagellate blooms (“red tides”). Environmental Science & Technology 52:5,718−5,724, https://doi.org/10.1021/acs.est.8b00335.
Armstrong, T., A.J. Khursigara, S.S. Killen, H. Fearnley, K.J. Parsons, and A.J. Esbaugh. 2019. Oil exposure alters social group cohesion in fish. Scientific Reports 9:1350, https://doi.org/10.1038/s41598-019-49994-1.
Barratclough, A., F.M. Gomez. J.S. Morey, A. Deming, C. Parry, J.M. Meegan, K.P. Carlin, L.H. Schwacke, S.K. Venn-Watson, E.D. Jensen, and C.R. Smith. 2020. Pregnancy profiles in the common bottlenose dolphin (Tursiops truncatus): Clinical biochemical and hematological variations during healthy gestation and a successful outcome. Theriogenology 142:92–103, https://doi.org/10.1016/j.theriogenology.2019.09.028.
Barron, M.G., S.C. Chiasson, and A.C. Bejarano. 2020. Ecotoxicology of deep ocean spills. Pp. 466–478 in Deep Oil Spills. S.A. Murawski, C.H. Ainsworth, S. Gilbert, D.J. Hollander, C.B. Paris, M. Schlüter, and D.L. Wetzel, eds, Springer Nature, https://doi.org/10.1007/978-3-030-11605-7.
Bautista, N.M., and W.W. Burggren. 2019. Parental stressor exposure simultaneously conveys both adaptive and maladaptive larval phenotypes through epigenetic inheritance in the zebrafish (Danio rerio). Journal of Experimental Biology 222:jeb208918, https://doi.org/10.1242/jeb.208918.
Bayha, K.M., N. Ortell, C.N. Ryan, K.J. Griffitt, M. Krasnec, J. Sena, T. Ramaraj, R. Takeshita, G.D. Mayer, F. Schilkey, and R.J. Griffitt. 2017. Crude oil impairs immune function and increases susceptibility to pathogenic bacteria in southern flounder. PLoS ONE 12:e0176559, https://doi.org/10.1371/journal.pone.0176559.
Bejarano, A.C., and M.G. Barron. 2014. Development and practical application of petroleum and dispersant interspecies correlation models for aquatic species. Environmental Science & Technology 48:4,564−4,572, https://doi.org/10.1021/es500649v.
Bejarano, A.C., J.R. Clark, and G.M. Coelho. 2014. Issues and challenges with oil toxicity data and implications for their use in decision making: A quantitative review. Environmental Toxicology and Chemistry 33:732–742, https://doi.org/10.1002/etc.2501.
Bentivegna, C.S., K.R. Cooper, G. Olson, E.A. Pena, D.R. Milleman, and R.J. Portier. 2015. Chemical and histological comparisons between Brevoortia sp. (menhaden) collected in fall 2010 from Barataria Bay, LA and Delaware Bay, NJ following the Deepwater Horizon (DWH) oil spill. Marine Environmental Research 112(Part A):21-34, https://doi.org/10.1016/j.marenvres.2015.08.011.
Beyer, J., H.C. Trannum, T. Bakke, P.V. Hodson, and T.K. Collier. 2016. Environmental effects of the Deepwater Horizon oil spill: A review. Marine Pollution Bulletin 110:28–51, https://doi.org/10.1016/j.marpolbul.2016.06.027.
Boesch, D.F., and N.N. Rabalais, eds. 1987. Long-term Environmental Effects of Offshore Oil and Gas Development. Elsevier Applied Science, New York, 708 pp., https://doi.org/10.4324/9780203497777.
Bonisoli-Alquati, A., W. Xu, P.C. Stouffer, and S.S. Taylor. 2020. Transcriptome analysis indicates a broad range of toxic effects of Deepwater Horizon oil on seaside sparrows. Science of the Total Environment 720:137583, https://doi.org/10.1016/j.scitotenv.2020.137583.
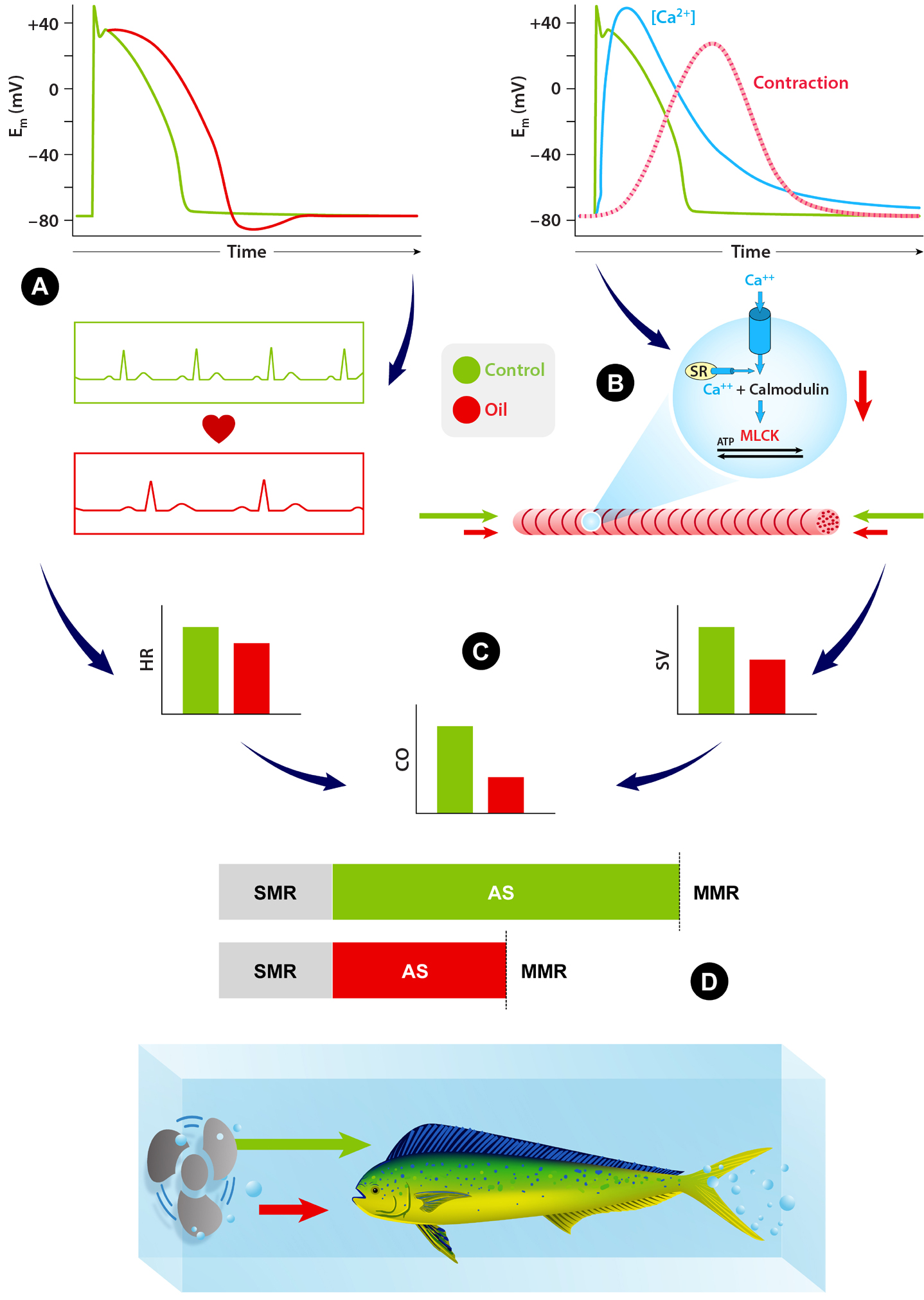
Brette, F., B. Machado, C. Cros, J.P. Incardona, N.L. Scholz, and B.A. Block. 2014. Crude oil impairs cardiac excitation-contraction coupling in fish. Science 343(6172):772–776, https://doi.org/10.1126/science.1242747.
Brette, F., H.A. Shiels, G.L.J. Galli, C. Cros, J.P. Incardona, N.L. Schloz, and B.A. Block. 2017. A novel cardiotoxic mechanism for a pervasive global pollutant. Scientific Reports 7:41476, https://doi.org/10.1038/srep41476.
Brewton, R.A., R. Fulford, and R.J. Griffitt. 2013. Gene expression and growth as indicators of effects of the BP Deepwater Horizon oil spill on spotted seatrout (Cynoscion nebulosus). Journal of Toxicology and Environmental Health, Part A 76(21):1,198–1,209, https://doi.org/10.1080/15287394.2013.848394.
Brown-Peterson, N.J., M. Krasnec, R. Takeshita, C.N. Ryan, K.J. Griffitt, C. Lay, G.D. Mayer, K.M. Bayha, W.E. Hawkins, I. Lipton, and others. 2015. A multiple endpoint analysis of the effects of chronic exposure to sediment contaminated with Deepwater Horizon oil on juvenile southern flounder and their associated microbiomes. Aquatic Toxicology 165:197–209, https://doi.org/10.1016/j.aquatox.2015.06.001.
Burns, K.A., and J.M. Teal. 1979. The West Falmouth oil spill: Hydrocarbons in the salt marsh ecosystem. Estuarine and Coastal Marine Science 8:349–360, https://doi.org/10.1016/0302-3524(79)90052-5.
Capuzzo, J.M. 1981. Predicting pollution effects in the marine environment. Oceanus 24:25–33.
Capuzzo, J.M. 1987. Biological effects of petroleum hydrocarbons: Assessments from experimental results. Pp. 343–410 in Long-term Environmental Effects of Offshore Oil and Gas Development. D.F. Boesch and N.N. Rabalais, eds, Elsevier Applied Science, London.
Collier, T.K., and U. Varanasi. 1987. Biochemical indicators of contaminant exposure in flatfish from Puget Sound, WA. Pp. 1,544–1,549 in Proceedings of Oceans ‘87 IEEE. Washington, DC, https://doi.org/10.1109/OCEANS.1987.1160572.
Cox, G.K., D.A. Crossley II, J.D. Stieglitz, R.M. Heuer, D.D. Benetti, and M. Grosell. 2017. Oil exposure impairs in situ cardiac function in response to β-adrenergic stimulation in cobia (Rachycentron canadum). Environmental Science & Technology 51:14,390–14,396, https://doi.org/10.1021/acs.est.7b03820.
Deak, K.L. 2020. A Health Evaluation of Gulf of Mexico Golden Tilefish (Lopholatilus chamaeleonticeps) and Red Snapper (Lutjanus campechanus) Following the Deepwater Horizon Oil Spill. PhD dissertation, College of Marine Science, University of South Florida, St. Petersburg, FL, 242 pp.
Deepwater Horizon Natural Resource Damage Assessment Trustees. 2016. Deepwater Horizon Oil Spill: Final Programmatic Damage Assessment and Restoration Plan and Final Programmatic Environmental Impact Statement, http://www.gulfspillrestoration.noaa.gov/restoration-planning/gulf-plan.
De Guise, S., M. Levin, L. Jasperse, J. Herrman, R.S. Wells, T. Rowles, and L. Schwacke. 2021. Long-term immunological alterations in bottlenose dolphin a decade after the Deepwater Horizon oil spill in the Northern Gulf of Mexico: Potential for multigenerational effects. Environmental Toxicology and Chemistry, https://doi.org/10.1002/etc.4980.
Deis, D.R., J.W. Fleeger, S.M. Bourgoin, I.A. Mendelssohn, Q. Lin, and A. Hou. 2017. Shoreline oiling effects and recovery of salt marsh macroinvertebrates from the Deepwater Horizon oil spill. PeerJ 5:e3680, https://doi.org/10.7717/peerj.3680.
Diamante, G., E.G. Xu, S. Chen, E. Mager, M. Grosell, and D. Schlenk. 2017. Differential expression of microRNAs in embryos and larvae of mahi-mahi (Coryphaena hippurus) exposed to Deepwater Horizon oil. Environmental Science & Technology Letters 4:523–529, https://doi.org/10.1021/acs.estlett.7b00484.
Dubansky, B., A. Whitehead, J.T. Miller, C.D. Rice, and F. Galvez. 2013. Multitissue molecular, genomic, and developmental effects of the deepwater horizon oil spill on resident gulf killifish (Fundulus grandis). Environmental Science & Technology 47:5,074–5,082, https://doi.org/10.1021/es400458p.
Esbaugh, A.J., E.M. Mager, J.D. Stieglitz, R. Hoenig, T.L. Brown, B.L. French, T.L. Linbo, H. Forth, N.L. Scholz, J.P. Incardona, and others. 2016. The effects of weathering and chemical dispersion on Deepwater Horizon crude oil toxicity to mahi-mahi (Coryphaena hippurus) early life stages. Science of the Total Environment 543(Part A):644–651, https://doi.org/10.1016/j.scitotenv.2015.11.068.
Fleeger, J.W., K.R. Carman, M.R. Riggio, I.A. Mendelssohn, Q. Lin, A. Hou, D.R. Deis, and S. Zengel. 2015. Recovery of salt marsh benthic microalgae and meiofauna following the Deepwater Horizon oil spill linked to recovery of Spartina alterniflora. Marine Ecology Progress Series 536:39–54, https://doi.org/10.3354/meps11451.
Fleeger, J.W., M.R. Riggio, I.A. Mendelssohn, Q. Lin, A. Hou, and D.R. Deis. 2018. Recovery of saltmarsh meiofauna six years after the Deepwater Horizon oil spill. Journal of Experimental Marine Biology and Ecology 502:182–190, https://doi.org/10.1016/j.jembe.2017.03.001.
Fleeger, J.W., M.R. Riggio, I.A. Mendelssohn, Q. Lin, D.R. Deis, D.S. Johnson, K.R. Carman, S.A. Graham, S. Zengel, and A. Hou. 2019. What promotes the recovery of salt marsh infauna after oil spills? Estuaries and Coasts 4:204–217, https://doi.org/10.1007/s12237-018-0443-2.
Fleeger, J.W., D.S. Johnson, S. Zengel, I.A. Mendelssohn, D.R. Deis, S.A. Graham, Q. Lin, M.C. Christman, M.R. Riggio, and M. Pant. 2020. Macroinfauna responses and recovery trajectories after an oil spill differ from those following saltmarsh restoration. Marine Environmental Research 161:105099, https://doi.org/10.1016/j.marenvres.2020.105099.
Fodrie, F.J., K.W. Able, F. Galvez, K.L. Heck Jr., O.P. Jensen, P.C. López-Duarte, C.W. Martin, R.E. Turner, and A. Whitehead. 2014. Integrating organismal and population responses of estuarine fishes in Macondo spill research. BioScience 64(9):778–788, https://doi.org/10.1093/biosci/biu123.
Frasier, K.E. 2020. Evaluating impacts of deep oil spills on oceanic marine mammals. Pp. 419–441 in Scenarios and Responses to Future Deep Oil Spills: Fighting the Next War. S.A. Murawski, C.H. Ainsworth, S. Gilbert, D.J. Hollander, C.B. Paris, M. Schlüter, and D.L. Wetzel, eds, Springer Nature, https://doi.org/10.1007/978-3-030-12963-7_25.
Frasier, K.E., A. Solsona-Berga, L. Stokes, and J.A. Hildebrand. 2020. Impacts of the Deepwater Horizon oil spill on marine mammals and sea turtles. Pp. 431–462 in Deep Oil Spills: Facts, Fate and Effects. S.A. Murawski, C.H. Ainsworth, S. Gilbert, D.J. Hollander, C.B. Paris, M. Schlüter, and D.L. Wetzel, eds, Springer Nature, https://doi.org/10.1007/978-3-030-11605-7.
Garcia, T.I., Y. Shen, D. Crawford, M.F. Oleksiak, A. Whitehead, and R.B. Walter. 2012. RNA-Seq reveals complex genetic response to Deepwater Horizon oil release in Fundulus grandis. BMC Genomics 13:474, https://doi.org/10.1186/1471-2164-13-474.
Garcia, S.M., K.T. Du Clos, O.H. Hawkins, and B.J. Gemmell. 2020. Sublethal effects of crude oil and chemical dispersants on multiple life history stages of the eastern oyster, Crassostrea virginica. Journal of Marine Science and Engineering 8:0808, https://doi.org/10.3390/jmse8100808.
Gomez, F.M., K.M. Colegrove, W.B. Musser, J.M. Meegan, R. Takeshita, A. Cárdenas Llerenas, T.K. Rowles, L.H. Schwacke, E.D. Jensen, and C.R. Smith. 2020. Potential mechanisms and associated predictors of pregnancy failure in bottlenose dolphins (Tursiops truncatus). Proceedings of the International Association of Aquatic Animal Medicine, Virtual Symposium hosted by the University of Florida.
Goodbody-Gringley, G., D.L. Wetzel, D. Gillon, E. Pulster, A. Miller, and K.B. Ritchie. 2013. Toxicity of Deepwater Horizon source oil and the chemical dispersant, Corexit® 9500, to coral larvae. PLoS ONE 8:e45574, https://doi.org/10.1371/journal.pone.0045574.
Grosell, M., R.J. Griffitt, T.A. Sherwood, and D.L. Wetzel. 2020. Digging deeper than LC/EC50: Nontraditional endpoints and non-model species in oil spill toxicology. Pp. 497–514 in Deep Oil Spills: Facts, Fate and Effects. S.A. Murawski, C.H. Ainsworth, S. Gilbert, D.J. Hollander, C.B. Paris, M. Schlüter, and D.L. Wetzel, eds, Springer Nature, https://doi.org/10.1007/978-3-030-11605-7_29.
Grosell, M., and C. Pasparakis. 2021. Physiological responses of fish to oil spills. Annual Reviews in Marine Science 13:137–160, https://doi.org/10.1146/annurev-marine-040120-094802.
Harrill, J., I. Shah, R.W. Setzer, D. Haggard, S. Auerbach, R. Judson, and R.S. Thomas. 2019. Considerations for strategic use of high-throughput transcriptomics chemical screening data in regulatory decisions. Current Opinions in Toxicology 15:64–75, https://doi.org/10.1016/j.cotox.2019.05.004.
Heuer, R.M., G. Galli, H. Shiels, L. Fieber, G. Cox, E. Mager, J. Stieglitz, D.D. Benetti, M. Grosell, and D. Crossley. 2019. Impacts of Deepwater Horizon crude oil on Mahi-Mahi (Coryphaena hippurus) heart cell function. Environmental Science and Technology 53(16):9,895–9,904, https://doi.org/10.1021/acs.est.9b03798.
Hollenbeck, C.M., D.S. Portnoy, D. Wetzel, T.A. Sherwood, P.B. Samollow, and J.R. Gold. 2017. Linkage mapping and comparative genomics of red drum (Sciaenops ocellatus) using next-generation sequencing. G3 Genes, Genomics, Genetics 7:843–850, https://doi.org/10.1534/g3.116.036350.
ICCOPR (Interagency Coordinating Committee on Oil Pollution Research). 2015. Oil Pollution Research and Technology Plan, Fiscal Years 2015–2021. 190 pp., https://www.dco.uscg.mil/Portals/9/CG-5R/ICCOPR/Files/2015-2021 Research and Technology Plan.pdf?ver=2018-01-05-133442-250.
Incardona, J.P., L.D. Gardner, T.L. Linbo, T.L. Brown, A.J. Esbaugh, E.M. Mager, J.D. Stieglitz, B.L. French, J.S. Labenia, C.A. Laetz, and others. 2014. Deepwater Horizon crude oil impacts the developing hearts of large predatory pelagic fish. Proceedings of the National Academy of Sciences of the United States of America 111(15):1,510–1,518, https://doi.org/10.1073/pnas.1320950111.
Incardona, J.P. 2017. Molecular mechanisms of crude oil developmental toxicity in fish. Archives in Environmental Contamination and Toxicology 73:19–32, https://doi.org/10.1007/s00244-017-0381-1.
Ivančić, M., F.M. Gomez, W.B. Musser, A. Barratclough,
J.M. Meegan, S.M. Waitt, A. Cárdenas Llerenas,
E.D. Jensen, and C.R. Smith. 2020. Ultrasonographic findings associated with normal pregnancy and fetal wellbeing in the bottlenose dolphin (Tursiops truncatus). Veterinary Radiology and Ultrasound 61:215–226, https://doi.org/10.1111/vru.12835.
Johansen, J.L., and A.J. Esbaugh. 2017. Sustained impairment of respiratory function and swim performance following acute oil exposure in a coastal marine fish. Aquatic Toxicology 187:82–89, https://doi.org/10.1016/j.aquatox.2017.04.002.
Kostka, J.E., S.B. Joye, and R.R. Colwell. 2020. Deepwater Horizon and the rise of the omics. EOS:101, https://doi.org/10.1029/2020EO140140.
Khursigara, A.J., P. Perrichon, N., Martinez Bautista, W.W. Burggren, and A.J. Esbaugh. 2017. Cardiac function and survival are affected by crude oil in larval red drum, Sciaenops ocellatus. Science of the Total Environment 579:797–804, https://doi.org/10.1016/j.scitotenv.2016.11.026.
Krahn, M.M., M.S. Myers, L.K. Moore, W.D. MacLeod Jr., and D.C. Malins. 1986. Associations between metabolites of aromatic compounds in bile and the occurrence of hepatic lesions in English sole (Parophrys vetulus) from Puget Sound, Washington. Archives of Environmental Contamination and Toxicology 15:61–67, https://doi.org/10.1007/BF01055249.
Lin, Q., and I.A. Mendelssohn. 2012. Impacts and recovery of the Deepwater Horizon oil spill on vegetation structure and function of coastal salt marshes in the northern Gulf of Mexico. Environmental Science & Technology 46(7):3,737–3,743, https://doi.org/10.1021/es203552p.
Lin, Q., I.A. Mendelssohn, S.A. Graham, A. Hou, J.W. Fleeger, and D.R. Deis. 2016. Response of salt marshes to oiling from the Deepwater Horizon spill: Implications for plant growth, soil surface-erosion, and shoreline stability. Science of the Total Environment 557–558:369–377, https://doi.org/10.1016/j.scitotenv.2016.03.049.
Linnehan, B.K., A. Hsu, F.M. Gomez, S.M. Huston, R. Takeshita, K.M. Colegrove, T.K. Rowles, A. Barratclough, W.B. Musser, C.A. Harms, and others. 2020. Standardization of dolphin cardiac auscultation and characterization of heart murmurs in managed and free-ranging bottlenose dolphins (Tursiops truncatus). Frontiers in Veterinary Science 7:570055, https://doi.org/10.3389/fvets.2020.570055.
Lubchenco, J., M.K. McNutt, G. Dreyfus, S.A. Murawski, D.M. Kennedy, P.T. Anastas, S. Chu, and T. Hunter. 2012. Science in support of the Deepwater Horizon response. Proceedings of the National Academy of Sciences of the United States of America 109:20,212–20,221, https://doi.org/10.1073/pnas.1204729109.
Mager, E.M., A.J. Esbaugh, J.D. Stieglitz, R. Hoenig, C. Bodinier, J.P. Incardona, N.L. Scholz, D.D. Benetti, and M. Grosell. 2014. Acute embryonic or juvenile exposure to Deepwater Horizon crude oil impairs the swimming performance of mahi-mahi (Coryphaena hippurus). Environmental Science & Technology 48:7,053−7,061, https://doi.org/10.1021/es501628k.
Meador, J.P., J.E. Stein, W.L. Reichert, and U. Varanasi. 1995. Bioaccumulation of polycyclic aromatic hydrocarbons by marine organisms. Pp. 79–171 in Reviews of Environmental Contamination and Toxicology. G. Ware, ed., Springer, New York, https://doi.org/10.1007/978-1-4612-2542-3_4.
Mitchelmore, C.L., A.C. Bejarano, and D.L. Wetzel. 2020. A synthesis of DWH oil: Chemical dispersant and chemically dispersed oil aquatic standard laboratory acute and chronic toxicity studies. Pp.480–496 in Deep Oil Spills: Facts, Fate and Effects. S.A. Murawski, C.H. Ainsworth, S. Gilbert, D.J. Hollander, C.B. Paris, M. Schlüter, and D.L. Wetzel, eds, Springer Nature, https://doi.org/10.1007/978-3-030-11605-7.
Morano, J.T., J.T. Tielens, C.A. Muirhead, B.J. Estabrook, P.J. Sullivan, P.J. Dugan, C.W. Clark, and A.N. Rice. 2020. Seasonal movements of Gulf of Mexico sperm whales following the Deepwater Horizon oil spill and the limitations of impact assessments. Marine Pollution Bulletin 161:111627, https://doi.org/10.1016/j.marpolbul.2020.111627.
Murawski, S.A., W.T. Hogarth, E.B. Peebles, and L. Barbieri. 2014. Prevalence of external skin lesions and polycyclic aromatic hydrocarbon concentrations in Gulf of Mexico fishes, post-Deepwater Horizon. Transactions of the American Fisheries Society 143(4):1,084–1,097, https://doi.org/10.1080/00028487.2014.911205.
Murawski, S.A., L. Hotaling, S. Gilbert, and D. Yoerger, eds. 2018. Advancing oil detection technologies for the marine ecosystem. Marine Technology Journal 52(6):7, https://doi.org/10.4031/MTSJ.52.6.3.
Murawski, S.A., D. Hollander, S. Gilbert, and A. Gracia. 2020. Deepwater oil and gas production in the Gulf of Mexico and related global trends. Pp. 16–32 in Scenarios and Responses to Future Deep Oil Spills: Fighting the Next War. S.A. Murawski, D. Hollander, C. Ainsworth, S. Gilbert, C.B. Paris, M. Schlüter, and D. Wetzel, eds, Springer Nature, https://doi.org/10.1007/978-3-030-12963-7.
Murawski, S.A., J.P. Kilborn, A.C. Bejarano, D. Chagaris, D. Donaldson, F.J. Hernandez Jr., T.C. MacDonald, C. Newton, E. Peebles, and K. Robinson 2021. A synthesis of Deepwater Horizon impacts on coastal and nearshore living marine resources. Frontiers in Marine Science 7:594862, https://doi.org/10.3389/fmars.2020.594862.
Myers, M.S., C.M. Stehr, O.P. Olson, L.L. Johnson, B.B. McCain, S.L. Chan, and U. Varanasi. 1994. Relationships between toxicopathic hepatic lesions and exposure to chemical contaminants in English sole (Pleuronectes vetulus), starry flounder (Platichthys stellatus), and white croaker (Genyonemus lineatus) from selected marine sites on the Pacific Coast, USA. Environmental Health Perspectives 102:200–215, https://doi.org/10.1289/ehp.94102200.
NASEM (National Academies of Science, Engineering and Medicine). 2020. The Use of Dispersants in Marine Oil Spill Response. The National Academies Press, Washington, DC, 340 pp., https://doi.org/10.17226/25161.
Nelson, D., R.M. Heuer, G.K. Cox, J.D. Stieglitz, R. Hoenig, E.M. Mager, D.D. Benetti, M. Grosell, and D.A. Crossley II. 2016. Effects of crude oil on in situ cardiac function in young adult mahi-mahi (Coryphaena hippurus). Aquatic Toxicology 180:274–81, https://doi.org/10.1016/j.aquatox.2016.10.012.
Nelson, D., J.D. Stieglitz, G.K. Cox, R.M. Heuer, D.D. Benetti, M. Grosell, and D.A. Crossley II. 2017. Cardio-respiratory function during exercise in the cobia, Rachycentron canadum: The impact of crude oil exposure. Comparative Biochemistry and Physiology C: Toxicology & Pharmacology 201:58–65, https://doi.org/10.1016/j.cbpc.2017.08.006.
NRC (National Research Council). 1985. Oil in the Sea: Inputs, Fates, and Effects. National Academies Press, Washington, DC, 581 pp., https://doi.org/10.17226/314.
Okey, A.B. 2007. An aryl hydrocarbon receptor odyssey to the shores of toxicology: The Deichmann Lecture, International Congress of Toxicology-XI. Toxicological Sciences 98:5–38, https://doi.org/10.1093/toxsci/kfm096.
Omar-Ali, A., C. Hohn, P.J. Allen, J. Rodriguez, and L. Petrie-Hanson. 2015. Tissue PAH, blood cell and tissue changes following exposure to water accommodated fractions of crude oil in alligator gar, Atractosteus spatula. Marine Environmental Research 108:33–44, https://doi.org/10.1016/j.marenvres.2015.04.011.
Park, B.S., D.L. Erdner, H.P. Bacosa, Z. Liu, and E.J. Buskey. 2020. Potential effects of bacterial communities on the formation of blooms of the harmful dinoflagellate Prorocentrum after the 2014 Texas City “Y” oil spill (USA). Harmful Algae 95:101802, https://doi.org/10.1016/j.hal.2020.101802.
Peacock, E.E., R.K. Nelson, A.R. Solow, J.D. Warren, J.L. Baker, and C.M. Reddy. 2005. The West Falmouth oil spill: ~100 kg of oil found to persist decades later. Environmental Forensics 6:273–281, https://doi.org/10.1080/15275920500194480.
Peiffer, R., and J. Cohen. 2015. Lethal and sublethal effects of oil, chemical dispersant, and dispersed oil on the ctenophore Mnemiopsis leidyi. Aquatic Biology 23:237–250, https://doi.org/10.3354/ab00625.
Perrichon, P., E.M. Mager, C. Pasparakis, J.D. Stieglitz, D.D. Benetti, M. Grosell, and W.W. Burggren. 2018. Combined effects of elevated temperature and Deepwater Horizon oil exposure on the cardiac performance of larval mahi-mahi, Coryphaena hippurus. PLoS ONE 13:e0203949, https://doi.org/10.1371/journal.pone.0203949.
Peterson, C.H., S.D. Rice, J.W. Short, D. Esler, J.L. Bodkin, B.E. Ballachey, and D.B. Irons. 2003. Long-term ecosystem response to the Exxon Valdez oil spill. Science 302:2,082–2,086, https://doi.org/10.1126/science.1084282.
Powers, S.P., C.H. Peterson, J. Cebrian, and K.L. Heck Jr. 2017. Response of nearshore ecosystems to the Deepwater Horizon spill. Marine Ecology Progress Series 576:107–110, https://doi.org/10.3354/meps12254.
Pulster, E.L., K. Main, D. Wetzel, and S. Murawski. 2017. Species-specific metabolism of naphthalene and phenanthrene in 3 species of marine teleosts exposed to Deepwater Horizon crude oil. Environmental Toxicology and Chemistry 36:3,168–3,176, https://doi.org/10.1002/etc.3898.
Pulster, E.L., A. Gracia, M. Armenteros, B.E. Carr, J. Mrowicki, and S.A. Murawski. 2020a. Chronic PAH exposures and associated declines in fish health indices observed for ten grouper species in the Gulf of Mexico. Science of the Total Environment 703:135551, https://doi.org/10.1016/j.scitotenv.2019.135551.
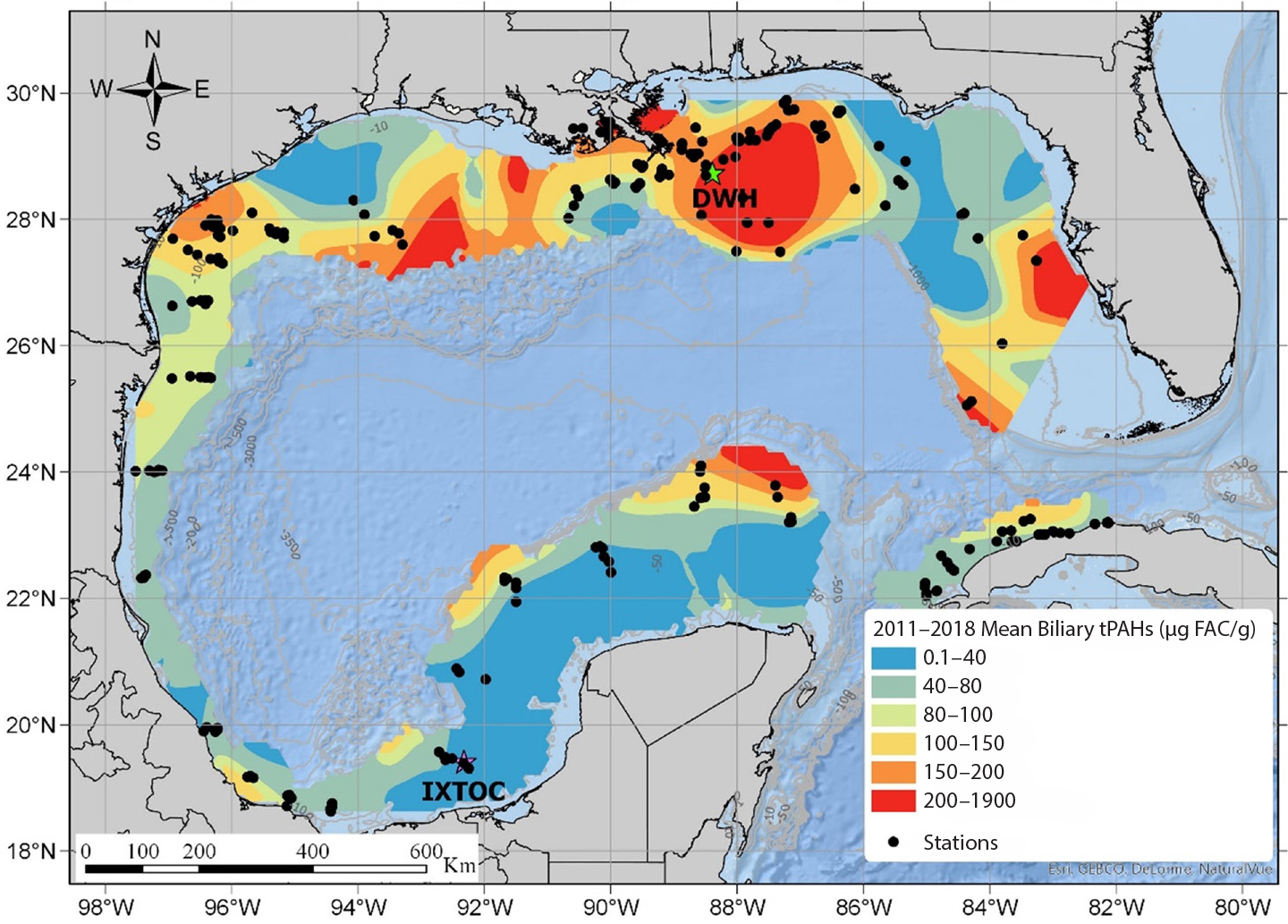
Pulster, E.L., A. Gracia, M. Armenteros, G. Toro-Farmer, S.M. Snyder, B.E. Carr, M.R. Schwaab, T.J. Nicholson, J. Mrowicki, and S.A. Murawski. 2020b. A first comprehensive baseline of hydrocarbon pollution in Gulf of Mexico fishes. Scientific Reports 10:6437, https://doi.org/10.1038/s41598-020-62944-6.
Pulster, E.L., A. Gracia, S.M. Snyder, K. Deak, S.B. Fogelson, and S.A. Murawski. 2020c. Chronic sub-lethal effects observed in wild-caught fishes following two major oil spills in the Gulf of Mexico: Deepwater Horizon and Ixoc 1. Pp. 388–413 in Deep Oil Spills: Facts, Fate and Effects. S.A. Murawski, C.H. Ainsworth, S. Gilbert, D.J. Hollander, C.B. Paris, M. Schlüter, and D.L. Wetzel, eds, Springer Nature, https://doi.org/10.1007/978-3-030-11605-7_24.
Pulster, E.L., A. Gracia, S.M. Snyder, I.C. Romero, B. Carr, G.A. Toro-Farmer, and S.A. Murawski. 2020d. Polycyclic aromatic hydrocarbon baselines in Gulf of Mexico fishes. Pp. 253–271 in Scenarios and Responses to Future Deep Oil Spills: Fighting the Next War. S.A. Murawski, C.H. Ainsworth, S. Gilbert, D.J. Hollander, C.B. Paris, M. Schlüter, and D.L. Wetzel, eds, Springer Nature, https://doi.org/10.1007/978-3-030-12963-7_15.
Pulster, E.L., S. Fogelson, B.E. Carr, J. Mrowicki, and S.A. Murawski. 2021. Hepatobiliary PAHs and prevalence of pathological changes in red snapper. Aquatic Toxicology 230:105714, https://doi.org/10.1016/j.aquatox.2020.105714.
Radović, J.R., I.C. Romero, T.B.P. Oldenburg, S.R. Larter, and J.W. Tunnell Jr. 2020. 40 years of weathering of coastal oil residues in the southern Gulf of Mexico. Pp. 328–340 in Deep Oil Spills: Facts, Fate and Effects. S.A. Murawski, C.H. Ainsworth, S. Gilbert, D.J. Hollander, C.B. Paris, M. Schlüter, and D.L. Wetzel, eds, Springer Nature, https://doi.org/10.1007/978-3-030-11605-7_20.
Reddy, C.M., T.L. Eglinton, A. Hounshell, H.K. White, L. Xu, R.B. Gaines, and G.G. Frysinger. 2002. The West Falmouth oil spill after thirty years: The persistence of petroleum hydrocarbons in marsh sediments. Environmental Science & Technology 36:4,754–4,760, https://doi.org/10.1021/es020656n.
Rice, S.D., R.B. Spies, D.A. Wolfe, and B.A. Wright, eds. 1996. Proceedings of the Exxon Valdez Oil Spill Symposium. American Fisheries Society, Bethesda, MD, 931 pp.
Robertson, M., A. Schrey, A. Shayter, C.J. Moss, and C. Richards. 2017. Genetic and epigenetic variation in Spartina alterniflora following the Deepwater Horizon oil spill. Evolutionary Applications 10:792–801, https://doi.org/10.1111/eva.12482.
Rodgers, M.L., T.A. Sherwood, A.M. Tarnecki, R.J. Griffitt, and D.L. Wetzel. 2021. Characterizing transcriptomic responses of southern flounder (Paralichthys lethostigma) chronically exposed to Deepwater Horizon oiled sediments. Aquatic Toxicology 230:105716, https://doi.org/10.1016/j.aquatox.2020.105716.
Romero, I.C., T.T. Sutton, B. Carr, E. Quintana-Rizzo, S.W. Ross, D.J. Hollander, and J.J. Torres. 2018. Decadal assessment of polycyclic aromatic hydrocarbons in mesopelagic fishes from the Gulf of Mexico reveals exposure to oil-derived sources. Environmental Science & Technology 52:10,985–10,996, https://doi.org/10.1021/acs.est.8b02243.
Romero, I.C., H. Judkins, and M. Vecchione. 2020. Temporal variability of polycyclic aromatic hydrocarbons in deep-sea cephalopods of the northern Gulf of Mexico. Frontiers in Marine Science 7:54, https://doi.org/10.3389/fmars.2020.00054.
Sanders, H.L., J.F. Grassle, G.R. Hampson, L.S. Morse, S. Garner-Price, and C.C. Jones. 1980. Anatomy of an oil spill: Long-term effects from the grounding of the barge Florida off West Falmouth, Massachusetts. Journal of Marine Research 38:265–380.
Schlenker, L.S., M.J. Welch, T.L. Meredith, E.M. Mager, E. Lari, E.A. Babcock, G.G. Pyle, P.L. Munday, and M. Grosell. 2019a. Damsels in distress: Oil exposure modifies behavior and olfaction in bicolor damselfish (Stegastes partitus). Environmental Science & Technology 53:10,993−11,001, https://doi.org/10.1021/acs.est.9b03915.
Schlenker, L.S., M.J. Welch, T.L. Meredith, E.M. Mager, J.D. Stieglitz, D.D. Benetti, P.L. Munday, and M. Grosell. 2019b. Exposure to crude oil from the Deepwater Horizon oil spill impairs oil avoidance behavior without affecting olfactory physiology in juvenile mahi-mahi (Coryphaena hippurus). Environmental Science & Technology 53:14,001–14,009, https://doi.org/10.1021/acs.est.9b05240.
Schwing, P.T., P.A. Montagna, S.B. Joye, C.B. Paris, E.E. Cordes, C.R. McClain, J.P. Kilborn, and S.A. Murawski. 2020. A synthesis of deep benthic faunal impacts and resilience following the Deepwater Horizon oil spill. Frontiers in Marine Science 7:560012, https://doi.org/10.3389/fmars.2020.560012.
Sherwood, T.A., R.L. Medvecky, C.A. Miller, A.M. Tarnecki, R.W. Schloesser, K.L. Main, C.L. Mitchelmore, and D.L. Wetzel. 2017. Nonlethal biomarkers of oxidative stress in oiled sediment exposed southern flounder (Paralichthys lethostigma): Utility for field-base monitoring exposure and potential recovery. Environmental Science & Technology 53:14,734–14,743, https://doi.org/10.1021/acs.est.9b05930.
Smith, C.R., T.K. Rowles, L.B. Hart, F.I. Townsend, R.S. Wells, E.S. Zolman, B.C. Balmer, B. Quigley, M. Ivančić, W. McKerche, and others. 2017. Slow recovery of Barataria Bay dolphin health following the Deepwater Horizon oil spill (2013–2014), with evidence of persistent lung disease and impaired stress response. Endangered Species Research 33:127–142, https://doi.org/10.3354/esr00778.
Smith, C.R., F.M. Gomez, K.M. Colegrove, T.K. Rowles, E.S. Zolman, T.R. Speakman, A. Barratclough, W.B. Musser, B.C. Balmer, R. Takeshita, and others. 2020. Sustained maternal illness and low reproductive success rate in Barataria Bay bottlenose dolphins (Tursiops truncatus) following the Deepwater Horizon oil spill. Proceedings of the International Association of Aquatic Animal Medicine. Virtual symposium hosted by the University of Florida.
Smith, C.R., T.K. Rowles, F.M. Gomez, K.M. Colegrove, R. Takeshita, E.S. Zolman, B.C. Balmer, R.S. Wells, F.I. Townsend, J.S. Morey, and others. 2021. Persistence of poor pulmonary health in dolphins living within the Deepwater Horizon oil spill footprint. Proceedings of the International Oil Spill Conference (virtual).
Snyder, S.M., E.L. Pulster, D.L. Wetzel, and S.A. Murawski. 2015. PAH exposure in Gulf of Mexico demersal fishes, post-Deepwater Horizon. Environmental Science & Technology 49(14):8,786–8,795, https://doi.org/10.1021/acs.est.5b01870.
Snyder, S.M., E.L. Pulster, and S.A. Murawski. 2019. Associations between chronic exposure to polycyclic aromatic hydrocarbons and health indices in Gulf of Mexico tilefish (Lopholatilus chamaeleonticeps) post Deepwater Horizon. Environmental Toxicology and Chemistry 38:2,659–2,671, https://doi.org/10.1002/etc.4583.
Snyder, S.M. 2020. Polycyclic Aromatic Hydrocarbon Exposure, Hepatic Accumulation, and Associated Microscopic Hepatic Changes in Gulf of Mexico Tilefish (Lopholatilus chamaeleonticeps). PhD dissertation, University of South Florida, College of Marine Science, 180 pp.
Snyder, S.M., J.A. Olin, E.L. Pulster, and S.A. Murawski. 2020. Spatial contrasts in hepatic and biliary PAHs in tilefish (Lopholatilus chamaeleonticeps) throughout the Gulf of Mexico, with comparison to the Northwest Atlantic. Environmental Pollution 258:113775, https://doi.org/10.1016/j.envpol.2019.113775.
Stegeman, J.J., J.J. Schlezinger, J.E. Craddock, and D.E. Tillitt. 2001. Cytochrome P450 1A expression in midwater fishes: Potential effects of chemical contaminants in remote oceanic zones. Environmental Science & Technology 35:54–62, https://doi.org/10.1021/es0012265.
Stein, J.E., W.L. Reichert, M. Nishimoto, and U. Varanasi. 1990. Overview of studies on liver carcinogenesis in English sole from Puget Sound; evidence for a xenobiotic chemical etiology II: Biochemical studies. Science of the Total Environment 94:51–69, https://doi.org/10.1016/0048-9697(90)90364-Z.
Stieglitz, J.D., E.M. Mager, R.H. Hoenig, D.D. Benetti, and M. Grosell. 2016a. Impacts of Deepwater Horizon crude oil exposure on adult mahi-mahi (Coryphaena hippurus) swim performance. Environmental Toxicology and Chemistry 35:2,613–2,622, https://doi.org/10.1002/etc.3436.
Stieglitz, J.D., E.M. Mager, R.H. Hoenig, M. Alloy, A.J. Esbaugh, C. Bodinier, D.D. Benetti, A.P. Roberts, and M. Grosell. 2016b. A novel system for embryo-larval toxicity testing of pelagic fish: Applications for impact assessment of Deepwater Horizon crude oil. Chemosphere 162:261–268, https://doi.org/10.1016/j.chemosphere.2016.07.069.
Sutton, T.T., T. Frank, H. Judkins, and I.C. Romero. 2020. As Gulf oil extraction goes deeper, who is at risk? Community structure, distribution, and connectivity of the deep-pelagic fauna. Pp. 403–418 in Scenarios and Responses to Future Deep Oil Spills. S.A. Murawski, C.H. Ainsworth, S. Gilbert, D.J. Hollander, C.B. Paris, M. Schlüter, and D.L. Wetzel, eds, Springer Nature, https://doi.org/
10.1007/978-3-030-12963-7_24.
Teal, J.M., J.W. Farrington, K.A. Burns, J.J. Stegeman, B.W. Tripp, B. Woodin, and C. Phinney. 1992. The West Falmouth oil spill after 20 years: Fate of fuel oil compounds and effects on animals. Marine Pollution Bulletin 24:607–614, https://doi.org/10.1016/0025-326X(92)90281-A.
Thomas, R.E., and S.D. Rice. 1987. Effect of water-soluble fraction of Cook Inlet crude oil on swimming performance and plasma cortisol in juvenile coho salmon (Oncorhynchus kisutch). Comparative Biochemistry and Physiology C 87:177–180, https://doi.org/10.1016/0742-8413(87)90200-3.
Transportation Research Board and National Research Council. 2003. Oil in the Sea III: Inputs, Fates, and Effects. National Academies Press, Washington, DC, 277 pp., https://doi.org/10.17226/10388.
Turner, N.R. 2020. Understanding the Toxicity of Single Hydrocarbons, Oil, and Dispersed Oil: A Species Sensitivity Assessment for Five Atlantic Coral Species. PhD dissertation, Nova Southeastern University, FL.
Turner, R.E., N.N. Rabalais, E.B. Overton, B.M. Meyer, G. McClenachan, E.M. Swenson, M. Besonen, M.L. Parsons, and J. Zingre. 2019. Oiling of the continental shelf and coastal marshes over eight years after the 2010 Deepwater Horizon oil spill. Environmental Pollution 252:1,367–1,376, https://doi.org/10.1016/j.envpol.2019.05.134.
Varanasi, U., D.J. Gmur, and P.A. Treseler. 1979. Influence of time and mode of exposure on biotransformation of naphthalene by juvenile starry flounder (Platichthys stellatus) and rock sole (Lepidopsetta bilineata). Archives of Environmental Contamination and Toxicology 8:673–692, https://doi.org/10.1007/BF01054869.
Varanasi, U., J.E. Stein, M. Nishimoto, W.L. Reichert, and T.K. Collier. 1987. Chemical carcinogenesis in feral fish: Uptake, activation, and detoxication of organic xenobiotics. Environmental Health Perspectives 71:155–170, https://doi.org/10.1289/ehp.8771155.
Wagner, H.-J. 2001. Sensory brain areas in mesopelagic fishes. Brain, Behavior and Evolution 57:117–133, https://doi.org/10.1159/000047231.
White, P.A., S. Robitaille, and J.B. Rasmussen. 1999. Heritable reproductive effects of Benzo[a]pyrene on the fathead minnow (Pimephales promelas). Environmental Toxicology and Chemistry 18:1,843–1,847, https://doi.org/10.1002/etc.5620180835.
Whitehead, A., B. Dubansky, C. Bodiner, T.I. Garcia, S. Miles, C. Pilley, V. Raghunathan, J.L. Roach, N. Walker, R.B. Walter, and others. 2012. Genomic and physiological footprint of the Deepwater Horizon oil spill on resident marsh fishes. Proceedings of the National Academy of Sciences of the United States of America 109(50):20,298–20,302, https://doi.org/10.1073/pnas.1109545108.
Whitehead, A., B.W. Clark, N.M. Reid, M.E. Hahn, and D. Nacci. 2017. When evolution is the solution to pollution: Key principles, and lessons from rapid repeated adaptation of killifish (Fundulus heteroclitus) populations. Evolutionary Applications 10:762–783, https://doi.org/10.1111/eva.12470.
Wiesenburg, D.A., B. Shipp, F.J. Fodrie, S. Powers, J. Lartigue, K.M. Darnell, M.M. Baustian, C. Ngo, J.F. Valentine, and K. Wowk. 2021. Prospects for Gulf of Mexico environmental recovery and restoration. Oceanography 34(1):164–173, https://doi.org/10.5670/oceanog.2021.124.
Xu, E.G., E.M. Mager, M. Grosell, C. Pasparakis, L.S. Schlenker, J.D. Stieglitz, D. Benetti, E.S. Hazard, S.M. Courtney, G. Diamante, and others. 2016. Time- and oil-dependent transcriptomic and physiological responses to Deepwater Horizon oil in mahi-mahi (Coryphaena hippurus) embryos and larvae. Environmental Science & Technology 50:7,842–7,851, https://doi.org/10.1021/acs.est.6b02205.
Xu, E.G., E.M. Mager, M. Grosell, E.S. Hazard, G. Hardiman, and D. Schlenk. 2017a. Novel transcriptome assembly and comparative toxicity pathway analysis in mahi-mahi (Coryphaena hippurus) embryos and larvae exposed to Deepwater Horizon oil. Scientific Reports 7:44546, https://doi.org/10.1038/srep44546.
Xu, E.G., A.J. Khursigara, J. Magnuson, E.S. Hazard, G. Hardiman, A.J. Esbaugh, A.P. Roberts, and D. Schlenk. 2017b. Larval red drum (Sciaenops ocellatus) sublethal exposure to weathered Deepwater Horizon crude oil: Developmental and transcriptomic consequences. Environmental Science & Technology 51:10,162–10,172, https://doi.org/10.1021/acs.est.7b02037.
Yednock, B.K., T.J. Sullivan, and J.E. Neigel. 2015. De novo assembly of a transcriptome from juvenile blue crabs (Callinectes sapidus) following exposure to surrogate Macondo crude oil. BMC Genomics 16:521, https://doi.org/10.1186/s12864-015-1739-2.
Zimmer, R.K., and C.A. Butman. 2000. Chemical signaling processes in the marine environment. Biological Bulletin 198:168–187, https://doi.org/10.2307/1542522.