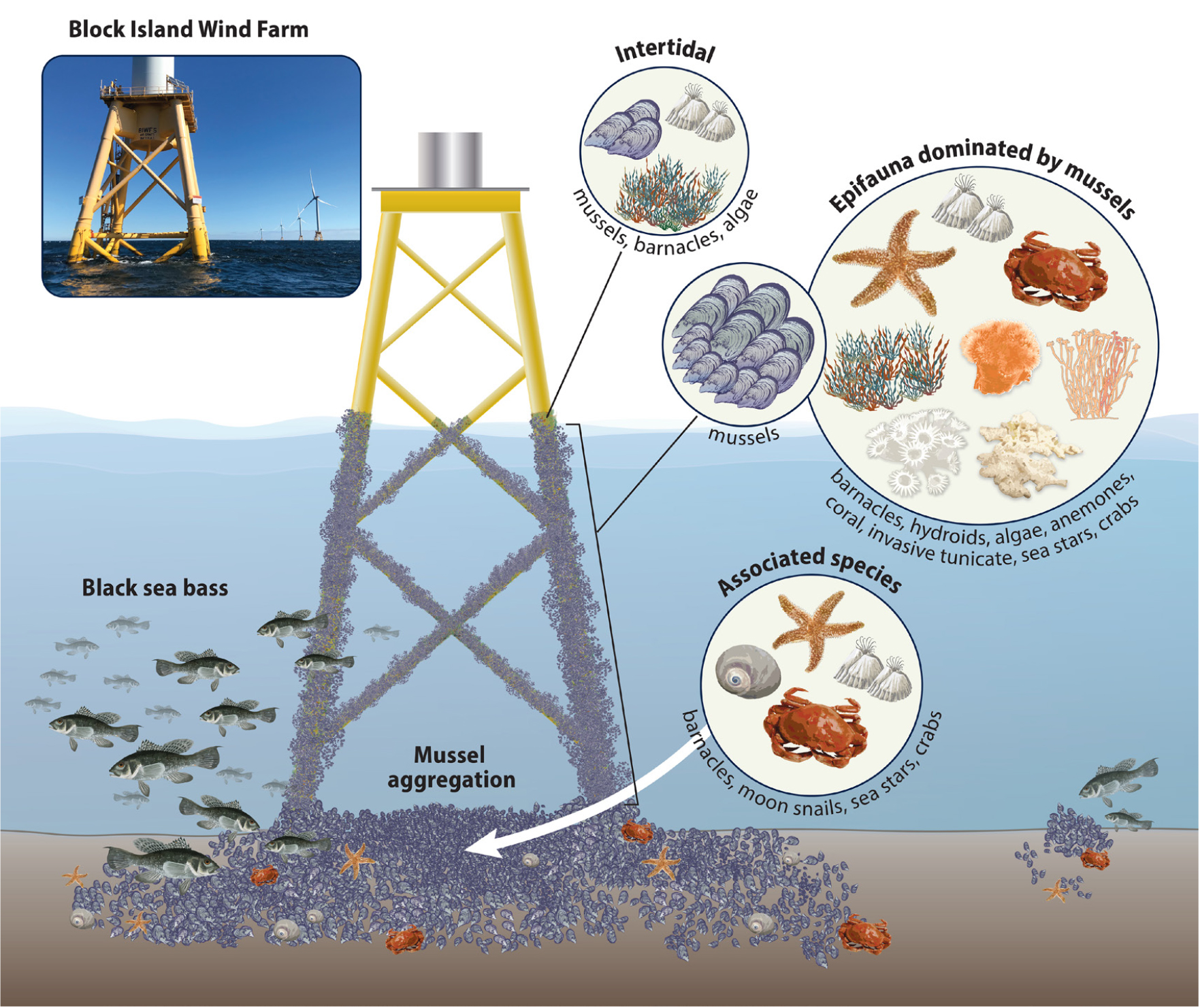
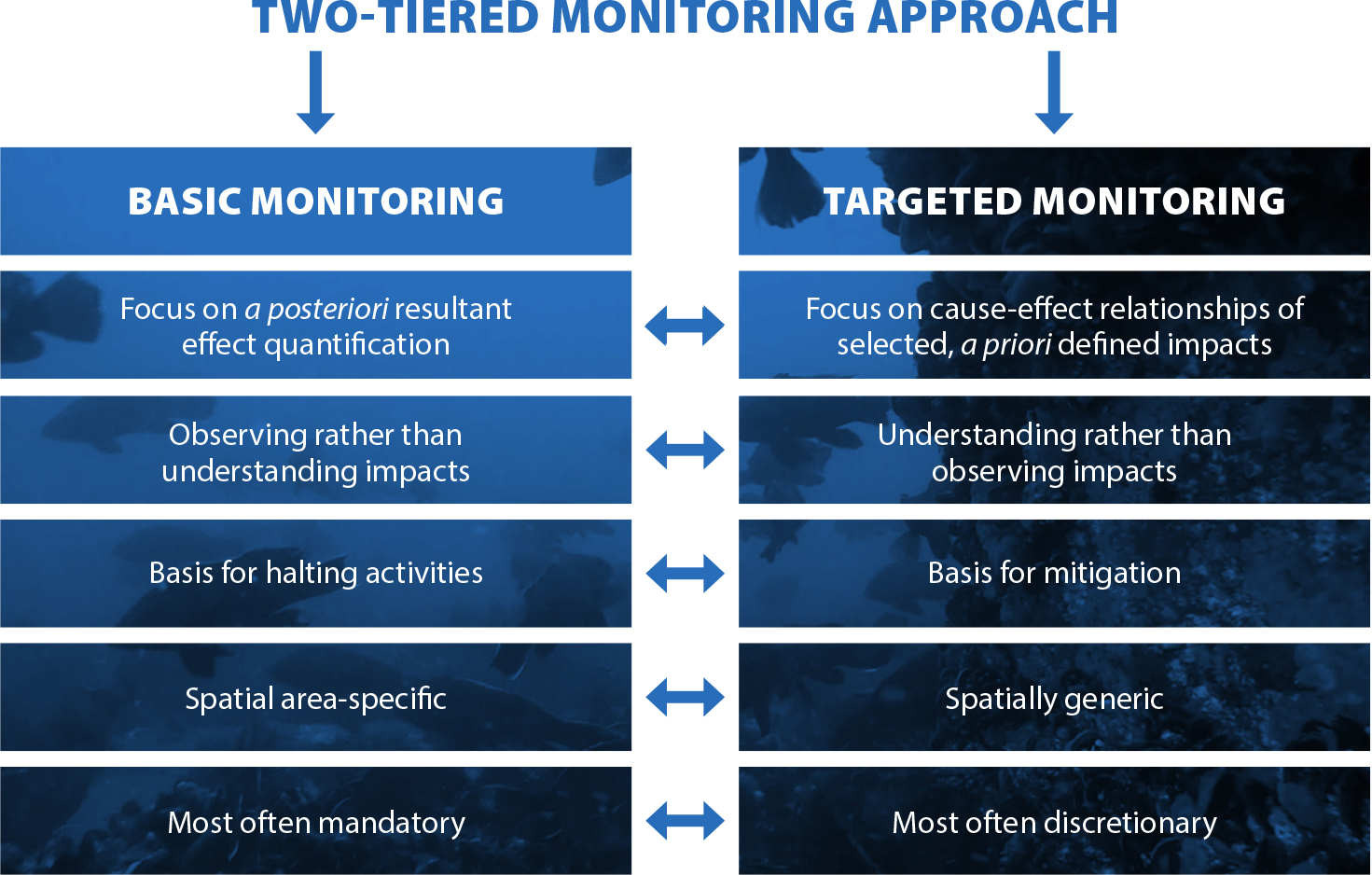
Bergström, L., F. Sundqvist, and U. Bergström. 2013. Effects of an offshore wind farm on temporal and spatial patterns in the demersal fish community. Marine Ecology Progress Series 485(1):199–210, https://doi.org/10.3354/meps10344.
BOEM (Bureau of Ocean Energy Management). 2019. Renewable Energy; Lease and Grant Information. https://www.boem.gov/renewable-energy/lease-and-grant-information.
BSH (Bundesamt fur Seeschifffahrt und Hydrographie). 2013. Standard Investigation of the Impacts of Offshore Wind Turbines on the Marine Environment (StUK4). BSH-Nr. 7003 for the German Federal Maritime and Hydrographic Agency, 87 pp.
Coates, D.A., Y. Deschutter, M. Vincx, and J. Vanaverbeke. 2014. Enrichment and shifts in macrobenthic assemblages in an offshore wind farm area in the Belgian part of the North Sea. Marine Environmental Research 95:1–12, https://doi.org/10.1016/j.marenvres.2013.12.008.
Coolen, J.W.P., A.R. Boon, R. Crooijmans, H. van Pelt, F. Kleissen, D. Gerla, J. Beermann, S.N.R. Birchenough, L.E. Becking, and P.C. Luttikhuizen. 2020. Marine stepping-stones: Connectivity of Mytilus edulis populations between offshore energy installations. Molecular Ecology 29(4):686–703, https://doi.org/10.1111/mec.15364.
CRMC (Coastal Resources Management Council). 2010. OceanSAMP, Volume 2. Rhode Island Ocean Special Area Management Plan, 2,922 pp., https://seagrant.gso.uri.edu/oceansamp/pdf/appendix/full_volume2_osamp_4.26.13.pdf.
Dannheim, J., L. Bergström, S.N.R. Birchenough, R. Brzana, A.R. Boon, J.W.P. Coolen, J.-C. Dauvin, I. De Mesel, J. Derweduwen, A.B. Gill, and others. 2020. Benthic effects of offshore renewables: Identification of knowledge gaps and urgently needed research. ICES Journal of Marine Science 77(3):1,092–1,108, https://doi.org/10.1093/icesjms/fsz018.
Degraer, S., R. Brabant, and Partnership. 2009. Recommendations for a future monitoring of wind farms in Belgium’s marine waters. Pp. 275–279 in Offshore Wind Farms in the Belgian Part of the North Sea: State of the Art After Two Years of Environmental Monitoring. S. Degraer and R. Brabant, eds, Royal Belgian Institute for Natural Sciences, Management Unit of the North Sea Mathematical Models, Marine Ecosystem Management Unit, Brussels.
Degraer, S., R. Brabant, B. Rumes, and L. Vigin, eds. 2019. Environmental Impacts of Offshore Wind Farms in the Belgian Part of the North Sea: Marking a Decade of Monitoring, Research and Innovation. Royal Belgian Institute of Natural Sciences, OD Natural Environment, Marine Ecology and Management, Brussels, 134 pp.
Degraer, S., D.A. Carey, J.W.P. Coolen, Z.L. Hutchison, F. Kerckhof, B. Rumes, and J. Vanaverbeke. 2020. Offshore wind farm artificial reefs affect ecosystem structure and functioning: A synthesis. Oceanography 33(4):48–57, https://doi.org/10.5670/oceanog.2020.405.
De Mesel, I., F. Kerckhof, A. Norro, B. Rumes, and S. Degraer. 2015. Succession and seasonal dynamics of the epifauna community on offshore wind farm foundations and their role as stepping stones for non-indigenous species. Hydrobiologia 756:37–50, https://doi.org/10.1007/s10750-014-2157-1.
Emu Limited. 2008. Kentish Flats Offshore Wind Farm Turbine Foundation Faunal Colonisation Diving Survey. Report No 08/J/1/03/1034/0839, Report by Vattenfall, 22 pp.
Fariñas-Franco, J.M., B. Pearce, J. Porter, D. Harries, J.M.M. Mair, A.S.S. Woolmer, W.G.G. Sanderson, J.M. Fariñas-Franco, B. Pearce, J. Porter, and others. 2014. Marine Strategy Framework Directive Indicators for Biogenic Reefs Formed by Modiolus modiolus, Mytius edulis and Sabellaria spinulosa: Part 1. Defining and Validating the Indicators. JNCC Report, No. 523, Heriot Watt University for JNCC, JNCC Peterborough, 379 pp.
FGDC (Federal Geographic Data Committee). 2012. Coastal and Marine Ecological Classification Standard. Marine and Coastal Spatial Data 1643 Subcommittee (Federal Geographic Data Committee). 353 pp.
Gilg, M.R., and T.J. Hilbish. 2003. The geography of marine larval dispersal: Coupling genetics with fine-scale physical oceanography. Ecology 84(11):2,989–2,998, https://doi.org/10.1890/02-0498.
Gill, A.B. 2005. Offshore renewable energy: Ecological implications of generating electricity in the coastal zone. Journal of Applied Ecology 42(4):605–615, https://doi.org/10.1111/j.1365-2664.2005.01060.x.
Grabowski, J.H., M. Bachman, C. Demarest, S. Eayrs, B.P. Harris, V. Malkoski, D. Packer, and D. Stevenson. 2014. Assessing the vulnerability of marine benthos to fishing gear impacts. Reviews in Fisheries Science & Aquaculture 22(2):142–155, https://doi.org/10.1080/10641262.2013.846292.
Grace, S. 2017. Winter quiescence, growth rate, and the release from competition in the temperate scleractinian coral Astrangia poculata (Ellis & Solander 1786). Northeastern Naturalist 24(sp7), https://doi.org/10.1656/045.024.s715.
Gubbay, S. 2007. Defining and Managing Sabellaria spinulosa reefs: Report of an Inter-Agency Workshop 1–2 May 2007. JNCC Report No. 405, JNCC, Peterborough, 26 pp.
GWEC (Global Wind Energy Council). 2019. Global Wind Report 2018. Global Wind Energy Council, Brussels, Belgium, 62 pp., https://gwec.net/wp-content/uploads/2019/04/GWEC-Global-Wind-Report-2018.pdf.
HDR. 2019. Field Observations During Wind Turbine Foundation Installation at the Block Island Wind Farm, Rhode Island. Final Report to the US Department of the Interior, Bureau of Ocean Energy Management, Office of Renewable Energy Programs, OCS Study BOEM 2019-028, 281 pp., https://espis.boem.gov/final reports/BOEM_2019-028.pdf.
HDR. 2020a. Benthic and Epifaunal Monitoring During Wind Turbine Installation and Operation at the Block Island Wind Farm, Rhode Island – Project Report. Final Report to the US Department of the Interior, Bureau of Ocean Energy Management, Office of Renewable Energy Programs, OCS Study BOEM 2020-044, Volume I, 263 pp; Volume II, 380 pp.
HDR. 2020b. Seafloor Disturbance and Recovery Monitoring at the Block Island Wind Farm, Rhode Island – Summary Report. Final Report to the US Department of the Interior, Bureau of Ocean Energy Management, Office of Renewable Energy Programs, OCS Study BOEM 2020-019, 63 pp.
Hendrick, V.J., and R.L. Foster-Smith. 2006. Sabellaria spinulosa reef: A scoring system for evaluating ‘reefiness’ in the context of the Habitats Directive. Journal of the Marine Biological Association of the United Kingdom 86(04):665–677, https://doi.org/10.1017/S0025315406013555.
Henriques, S., M.P. Pais, R.P. Vasconcelos, A. Murta, M. Azevedo, M.J. Costa, and H.N. Cabral. 2014. Structural and functional trends indicate fishing pressure on marine fish assemblages. Journal of Applied Ecology 51(3):623–631, https://doi.org/10.1111/1365-2664.12235.
Hutchings, P. 1998. Biodiversity and functioning of polychaetes in benthic sediments. Biodiversity & Conservation 7(9):1,133–1,145, https://doi.org/10.1023/A:1008871430178.
Jacobson, L.D. 2005. Longfin Inshore Squid, Loligo pealeii, Life History and Habitat Characteristics, Second Edition. NOAA Technical Memorandum NMFS-NE-193, US Department of Commerce, National Marine Fisheries Service, 52 pp.
Kerckhof, F., B. Rumes, T. Jacques, S. Degraer, and A. Noro. 2010. Early development of the subtidal marine biofouling on a concrete offshore windmill foundation on the Thornton Bank (southern North Sea): First monitoring results. International Journal of the Society for Underwater Technology 29(3):137–149, https://doi.org/10.3723/ut.29.137.
Kerckhof, F., B. Rumes, and S. Degraer. 2017. On the replicability of natural gravel beds by artificial hard substrata in Belgian waters. Pp. 73–84 in Environmental Impacts of Offshore Wind Farms in the Belgian Part of the North Sea: A Continued Move Towards Integration and Quantification. S. Degraer, R. Brabant, B. Rumes, and L. Vigin, eds, Royal Belgian Institute of Natural Sciences, OD Natural Environment, Marine Ecology and Management Section, Brussels.
Kritzer, J.P., M.B. DeLucia, E. Greene, C. Shumway, M.F. Topolski, J. Thomas-Blate, L.A. Chiarella, K.B. Davy, and K. Smith. 2016. The importance of benthic habitats for coastal fisheries. BioScience 66(4):274–284, https://doi.org/10.1093/biosci/biw014.
Krone, R., L. Gutow, T.J. Joschko, and A. Schröder. 2013. Epifauna dynamics at an offshore foundation – Implications of future wind power farming in the North Sea. Marine Environmental Research 85:1–12, https://doi.org/10.1016/j.marenvres.2012.12.004.
Krone, R., G. Dederer, P. Kanstinger, P. Krämer, C. Schneider, and I. Schmalenbach. 2017. Mobile demersal megafauna at common offshore wind turbine foundations in the German Bight (North Sea) two years after deployment – Increased production rate of Cancer pagurus. Marine Environmental Research 123:53–61, https://doi.org/10.1016/j.marenvres.2016.11.011.
Lefaible, N., L. Colson, U. Braeckman, and T. Moens. 2019. Evaluation of turbine-related impacts on macrobenthic communities withing two offshore wind farms during the operational phase. Pp. 47–64 in Environmental Impacts of Offshore Wind Farms in the Belgian Part of the North Sea: Marking a Decade of Monitoring, Research and Innovation. S. Degraer, R. Brabant, B. Rumes, and L. Vigin, eds, Royal Belgian Institute of Natural Sciences, OD Natural Environment, Marine Ecology and Management, Brussels.
LaFrance, M., J.W. King, B.A. Oakley, and S. Pratt. 2014. A comparison of top-down and bottom-up approaches to benthic habitat mapping to inform offshore wind energy development. Continental Shelf Research 83:24–44, https://doi.org/10.1016/j.csr.2014.04.007.
Lindeboom, H., S. Degraer, J. Dannheim, A.B. Gill, and D. Wilhelmsson. 2015. Offshore wind park monitoring programmes, lessons learned and recommendations for the future. Hydrobiologia 756(1):169–180, https://doi.org/10.1007/s10750-015-2267-4.
Maar, M., K. Bolding, J.K. Petersen, J.L.S. Hansen, and K. Timmermann. 2009. Local effects of blue mussels around turbine foundations in an ecosystem model of Nysted off-shore wind farm, Denmark. Journal of Sea Research 62(2):159–174, https://doi.org/10.1016/j.seares.2009.01.008.
Malek, A., M. LaFrance, J. Collie, and J. King. 2010. Fisheries ecology in Rhode Island and Block Island Sounds for the Rhode Island Ocean Special Area Management Plan 2010. Pp. 2,102–2,158 in oceanSAMP, Volume 2: Rhode Island Ocean Special Area Management Plan, Coastal Resources Management Council.
Manoukian, S., A. Spagnolo, G. Scarcella, E. Punzo, R. Angelini, and G. Fabi. 2010. Effects of two offshore gas platforms on soft-bottom benthic communities (northwestern Adriatic Sea, Italy). Marine Environmental Research 70(5):402–410, https://doi.org/10.1016/j.marenvres.2010.08.004.
Mavraki, N. 2020. On the Food-Web Ecology of Offshore Wind Farms, the Kingdom of Suspension Feeders. Doctoral dissertation, Ghent University, 278 pp.
Mavraki, N., S. Degraer, J. Vanaverbeke, and U. Braeckman. 2020. Organic matter assimilation by hard substrate fauna in an offshore wind farm area: A pulse-chase study. ICES Journal of Marine Science fsaa133, https://doi.org/10.1093/icesjms/fsaa133.
Methratta, E.T., and W.R. Dardick. 2019. Meta-analysis of finfish abundance at offshore wind farms. Reviews in Fisheries Science & Aquaculture 27(2):242–260, https://doi.org/10.1080/23308249.2019.1584601.
Nixon, S., S. Granger, C. Oviatt, L. Fields, and J. Mercer. 2010. Spatial and temporal variability of surface chlorophyll, primary production, and benthic metabolism in Rhode Island and Block Island Sounds. Pp. 652–704 in oceanSAMP, Volume 2: Rhode Island Ocean Special Area Management Plan. Coastal Resources Management Council.
Page, H.M., J.E. Dugan, D.S. Dugan, J.B. Richards, and D.M. Hubbard. 1999. Effects of an offshore oil platform on the distribution and abundance of commercially important crab species. Marine Ecology Progress Series 185:47–57, https://doi.org/10.3354/meps185047.
Reubens, J.T., S. Degraer, and M. Vincx. 2014. The ecology of benthopelagic fishes at offshore wind farms: A synthesis of 4 years of research. Hydrobiologia 727:121–136, https://doi.org/10.1007/s10750-013-1793-1.
Riisgård, H.U.S., P.P. Egede, I.B. Saavedra, and I. Barreiro Saavedra. 2011. Feeding behaviour of the mussel, Mytilus edulis: New observations, with a minireview of current knowledge. Journal of Marine Biology 2011:312459, https://doi.org/10.1155/2011/312459.
Schröder, A., C. Orejas, and T. Joschko. 2006. Benthos in the vicinity of piles: FINO 1 (North Sea). Pp. 185–200 in Offshore Wind Energy: Research on Environmental Impacts. J. Köller, J. Köppel, and W. Peters, eds, Springer Berlin Heidelberg, https://doi.org/10.1007/978-3-540-34677-7_12.
Schwartzbach, A., J. Behrens, and J. Svendsen. 2020. Atlantic cod Gadus morhua save energy on stone reefs: Implications for the attraction versus production debate in relation to reefs. Marine Ecology Progress Series 635:81–87, https://doi.org/10.3354/meps13192.
Slavik, K., C. Lemmen, W. Zhang, O. Kerimoglu, K. Klingbeil, and K.W. Wirtz. 2019. The large-scale impact of offshore wind farm structures on pelagic primary productivity in the southern North Sea. Hydrobiologia 845(1):35–53, https://doi.org/10.1007/s10750-018-3653-5.
Steimle, F.W. 1982. The benthic macroinvertebrates of the Block Island Sound. Estuarine, Coastal and Shelf Science 15(1):1–16, https://doi.org/10.1016/0272-7714(82)90032-4.
ten Brink, T.S., and T. Dalton. 2018. Perceptions of commercial and recreational fishers on the potential ecological impacts of the Block Island Wind Farm (US). Frontiers in Marine Science 5(439):1–13, https://doi.org/10.3389/fmars.2018.00439.
Trager, G.C., J.-S. Hwang, and J.R. Strickler. 1990. Barnacle suspension-feeding in variable flow. Marine Biology 105(1):117–127, https://doi.org/10.1007/BF01344277.
UNECE (United Nations Economic Commission for Europe). 1998. The Aarhus Convention.
Valentine, P.C., M.R. Carman, J. Dijkstra, and D.S. Blackwood. 2009. Larval recruitment of the invasive colonial ascidian Didemnum vexillum, seasonal water temperatures in New England coastal and offshore waters, and implications for spread of the species. Aquatic Invasions 4(1):153–168, https://doi.org/10.3391/ai.2009.4.1.16.
Wilber, D., L. Read, M. Griffin, and D. Carey. 2020. Block Island Wind Farm Demersal Fish Trawl Survey Synthesis Report – Years 1 to 6, October 2012 through September 2018. 182 pp.
Wilding, T.A., A.B. Gill, A. Boon, E. Sheehan, J.C. Dauvin, J.-P.P. Pezy, F. O’Beirn, U. Janas, L. Rostin, I. De Mesel, and others. 2017. Turning off the DRIP (‘Data-rich, information-poor’) – Rationalising monitoring with a focus on marine renewable energy developments and the benthos. Renewable and Sustainable Energy Reviews 74:848–859, https://doi.org/10.1016/j.rser.2017.03.013.
Wilhelmsson, D., T. Malm, and M.C. Öhman. 2006. The influence of offshore windpower on demersal fish. ICES Journal of Marine Science 63(5):775–784, https://doi.org/10.1016/j.icesjms.2006.02.001.
Willsteed, E., A.B. Gill, S.N.R. Birchenough, and S. Jude. 2017. Assessing the cumulative environmental effects of marine renewable energy developments: Establishing common ground. Science of the Total Environment 577:19–32, https://doi.org/10.1016/j.scitotenv.2016.10.152.
Wolfson, A., G. Van Blaricom, N. Davis, and G.S. Lewbel. 1979. The marine life of an offshore oil platform. Marine Ecology Progress Series 1:81–89, https://doi.org/10.3354/meps001081.
WOZEP (Dutch Governmental Offshore Wind Ecological Programme). 2016. Offshore wind energy ecological programme (Wozep); Monitoring and research programme 2017–2021. Final Report, Rijkswaterstaat, Ministry of Infrastructure and the Environment, 69 pp.
Zemeckis, D.R., D. Martins, L.A. Kerr, and S.X. Cadrin. 2014. Stock identification of Atlantic cod (Gadus morhua) in US waters: An interdisciplinary approach. ICES Journal of Marine Science 71(6):1,490–1,506, https://doi.org/10.1093/icesjms/fsu032.