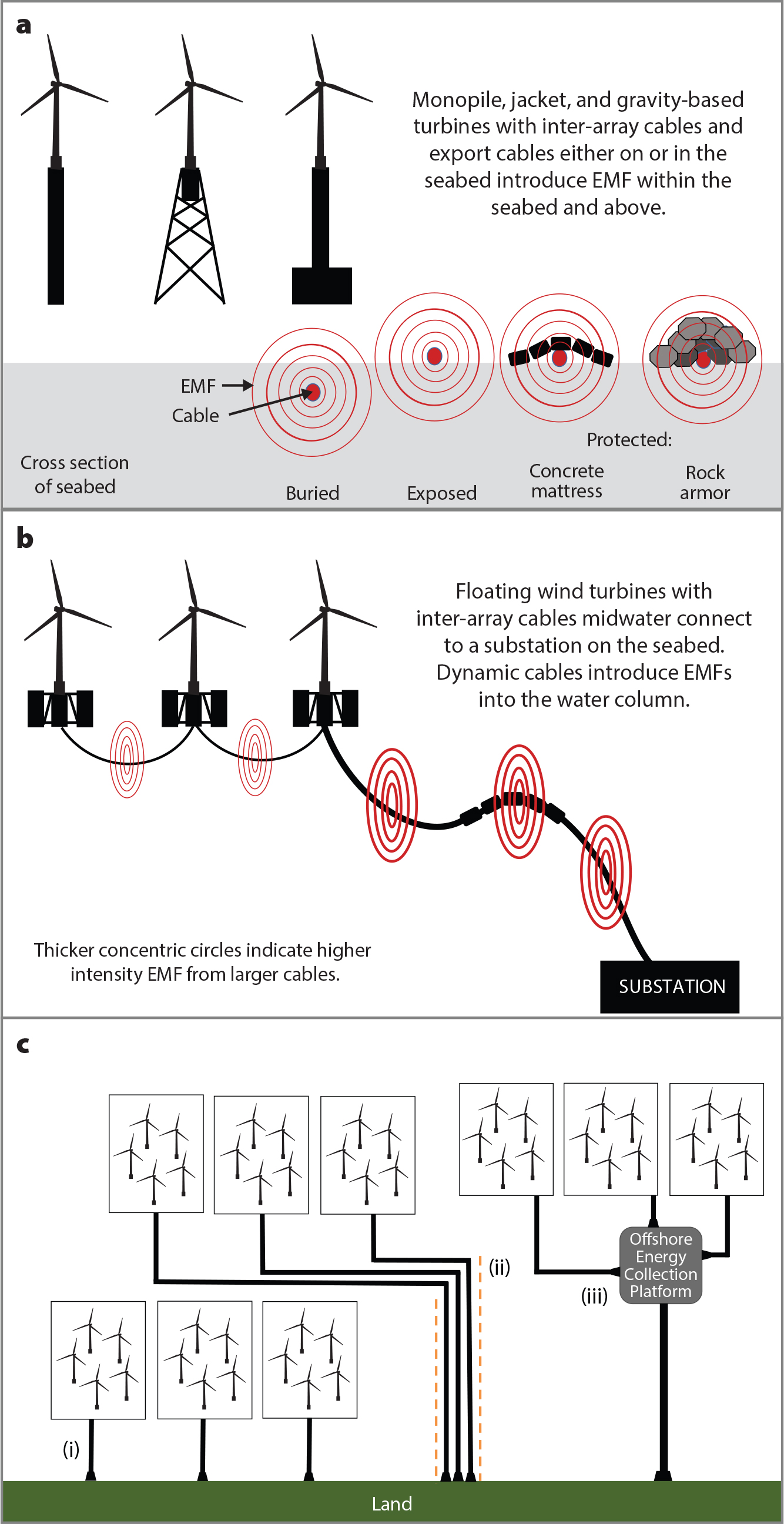
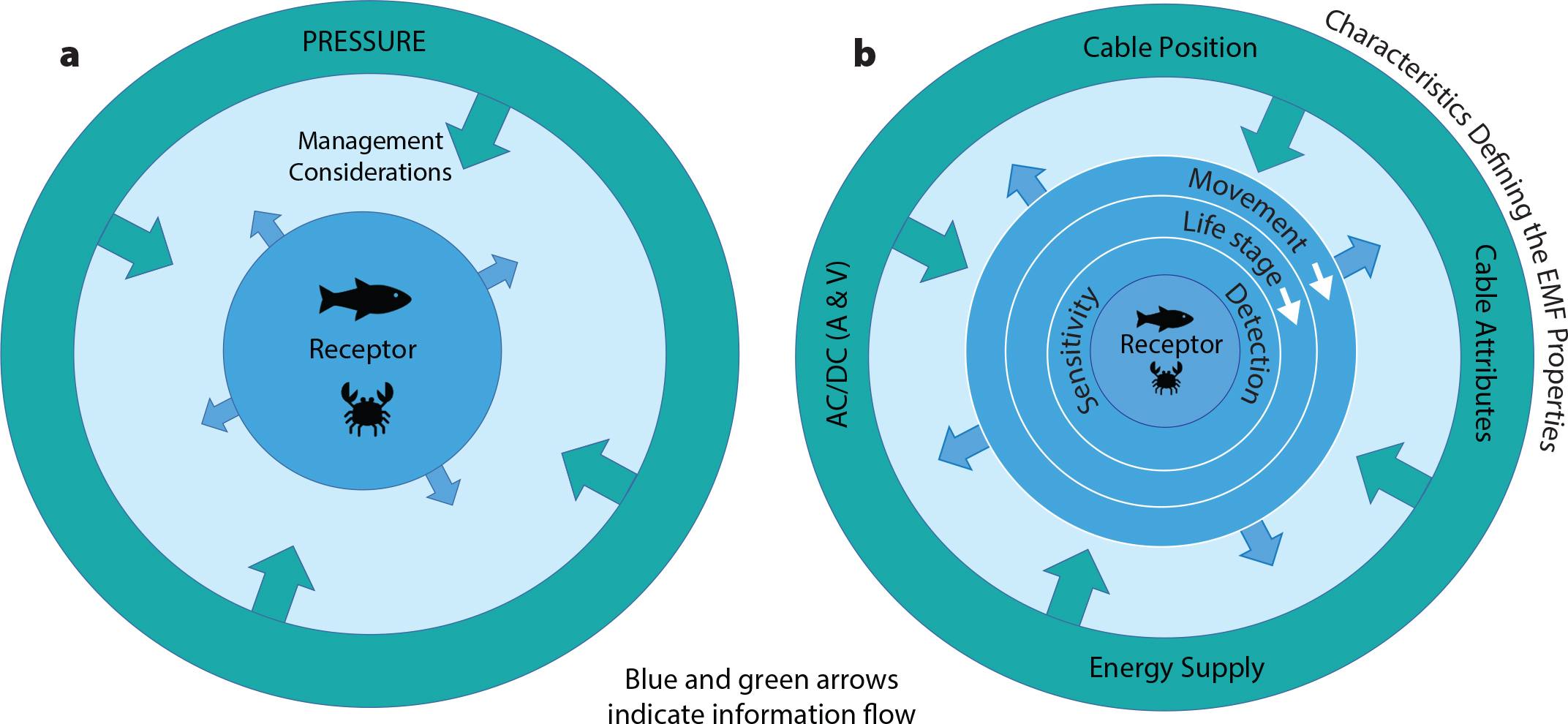
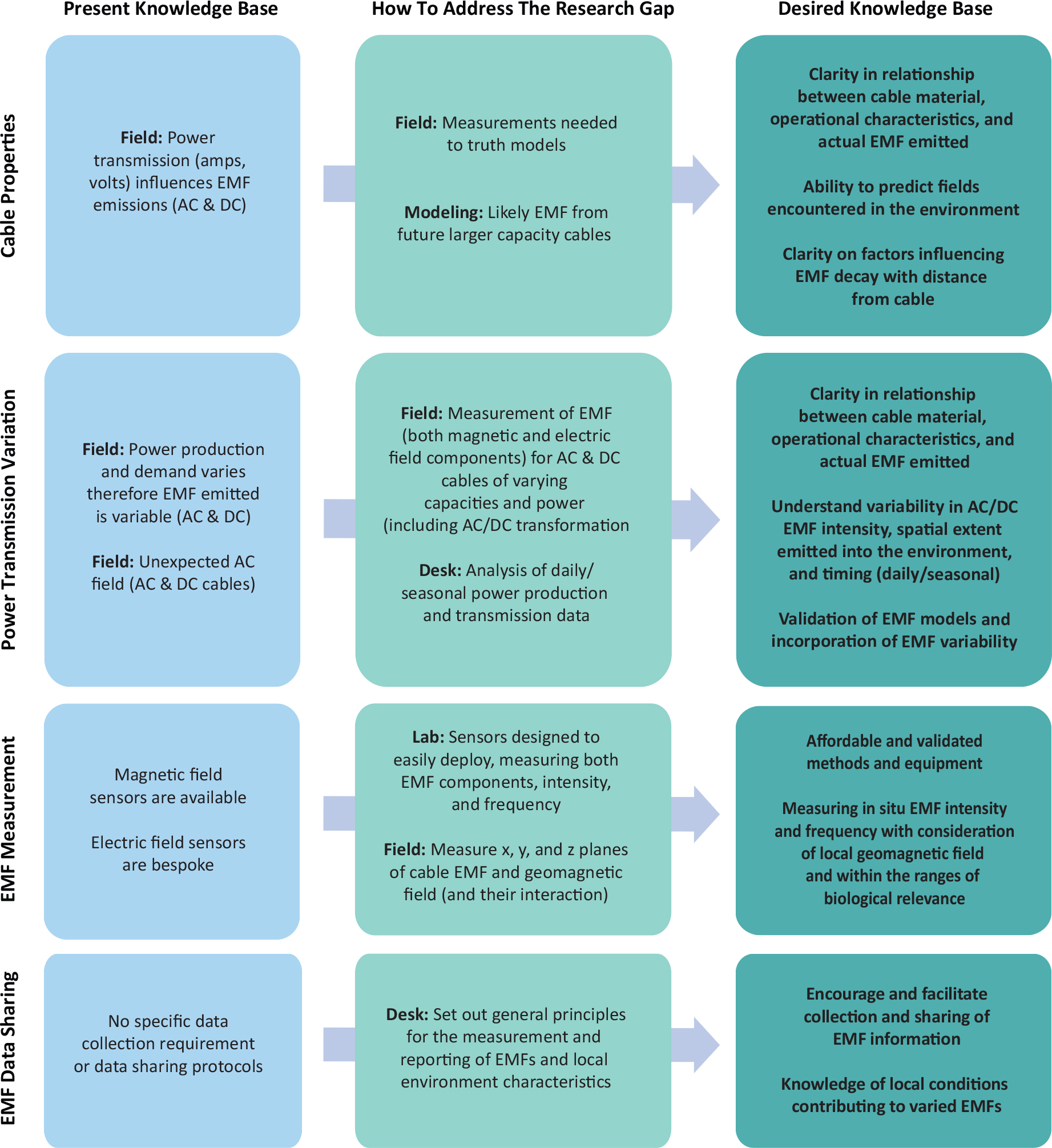
Anderson, J.M., T.M. Clegg, L.V.M.V.Q. Véras, and K.N. Holland. 2017. Insight into shark magnetic field perception from empirical observations. Scientific Reports 7(1):11042, https://doi.org/10.1038/s41598-017-11459-8.
Baker, C.V.H., M.S. Modrell, and J.A. Gillis. 2013. The evolution and development of vertebrate lateral line electroreceptors. The Journal of Experimental Biology 216(13):2,515–2,522, https://doi.org/10.1242/jeb.082362.
Ball, R.E., M.K. Oliver, and A.B. Gill. 2016. Early life sensory ability-ventilatory responses of thornback ray embryos (Raja clavata) to predator-type electric fields. Developmental Neurobiology 76(7):721–729, https://doi.org/10.1002/dneu.22355.
Bedore, C.N., and S.M. Kajiura. 2013. Bioelectric fields of marine organisms: Voltage and frequency contributions to detectability by electroreceptive predators. Physiological and Biochemical Zoology 86(3):298–311, https://doi.org/10.1086/669973.
Bochert, R., and M.L. Zettler. 2004. Long-term exposure of several marine benthic animals to static magnetic fields. Bioelectromagnetics 25(7):498–502, https://doi.org/10.1002/bem.20019.
Boehlert, G.W., and A.B. Gill. 2010. Environmental and ecological effects of ocean renewable energy development: A current synthesis. Oceanography 23(2):68–81, https://doi.org/10.5670/oceanog.2010.46.
Boles, L.C., and K.J. Lohmann. 2003. True navigation and magnetic maps in spiny lobsters. Nature 421(6918):60–63, https://doi.org/10.1038/nature01226.
Chase, B.C. 2002. Differences in diet of Atlantic bluefin tuna (Thunnus thynnus) at five seasonal feeding grounds on the New England continental shelf. Fishery Bulletin 100(2):168–180, https://spo.nmfs.noaa.gov/sites/default/files/pdf-content/2002/1002/chasef.pdf.
CMACS (Centre for Marine and Coastal Studies). 2003. A Baseline Assessment of Electromagnetic Field Generated by Offshore Windfarm Cables. University of Liverpool, 66 pp.
Corten, A. 2002. The role of “conservatism” in herring migrations. Reviews in Fish Biology and Fisheries 11(4):339–361, https://doi.org/10.1023/A:1021347630813.
Cresci, A., C.B. Paris, C.M.F. Durif, S. Shema, R.M. Bjelland, A.B. Skiftesvik, and H.I. Browman. 2017. Glass eels Anguilla anguilla have a magnetic compass linked to the tidal cycle. Science Advances 3(6):9, https://doi.org/10.1126/sciadv.1602007.
Dean, M.J., W.S. Hoffman, D.R. Zemeckis, and M.P. Armstrong. 2014. Fine-scale diel and gender-based patterns in behaviour of Atlantic cod (Gadus morhua) on a spawning ground in the Western Gulf of Maine. ICES Journal of Marine Science71(6):1,474–1,489, https://doi.org/10.1093/icesjms/fsu040.
Degraer, S., D.A. Carey, J.W.P. Coolen, Z.L. Hutchison, F. Kerckhof, B. Rumes, and J. Vanaverbeke. 2020. Offshore wind farm artificial reefs affect ecosystem structure and functioning: A synthesis. Oceanography 33(4):48–57, https://doi.org/10.5670/oceanog.2020.405.
Dhanak, M., E. An, R. Coulson, J. Frankenfield, S. Ravenna, D. Pugsley, G. Valdes, and W. Venezia. 2015. AUV-based characterization of EMF emissions from submerged power cables. Pp. 1–6 in MTS/IEEE OCEANS 2015: Discovering Sustainable Ocean Energy for a New World, https://doi.org/10.1109/OCEANS-Genova.2015.7271719.
Durif, C.M.F., H.I. Browman, J.B. Phillips, A.B. Skiftesvik, L.A. Vøllestad, and H.H. Stockhausen. 2013. Magnetic compass orientation in the European eel. PLoS ONE 8(3):e59212, https://doi.org/10.1371/journal.pone.0059212.
Fluharty, D. 2000. Habitat protection, ecological issues, and implementation of the sustainable fisheries act. Ecological Applications 10(2):325–337, https://doi.org/10.1890/1051-0761(2000)010[0325:HPEIAI]2.0.CO;2.
Formicki, K., A. Korzelecka-Orkisz, and A. Tański. 2019. Magnetoreception in fish. Journal of Fish Biology 95(1;Special Issue): 73–91, https://doi.org/10.1111/jfb.13998.
Franco, A., V. Quintino, and M. Elliott. 2015. Benthic monitoring and sampling design and effort to detect spatial changes: A case study using data from offshore wind farm sites. Ecological Indicators 57:298–304, https://doi.org/10.1016/j.ecolind.2015.04.040.
Gauldie, R.W., and G.D. Sharp. 1996. Skipjack velocity, dwell time and migration. Fisheries Oceanography 5(2):100–113, https://doi.org/10.1111/j.1365-2419.1996.tb00109.x.
Gill, A.B., Y. Huang, I. Gloyne-Phillips, J. Metcalfe, V. Quayle, J. Spencer, and V. Wearmouth. 2009. COWRIE 2.0 Electromagnetic Fields (EMF) Phase 2: EMF-Sensitive Fish Response to EM Emissions from Subsea Electricity Cables of the Type Used by the Offshore Renewable Energy Industry. Commissioned by COWRIE Ltd (project reference COWRIE-EMF-1-06), 128 pp.
Gill, A.B., Y. Huang, J. Spencer, and I. Gloyne-Philips. 2012. Electromagnetic Fields Emitted by High Voltage Alternating Current Offshore Wind Power Cables and Interactions with Marine Organisms. Institute of Engineering and Technology, London, 5 pp.
Gill, A.B., I. Gloyne-Philips, J. Kimber, and P. Sigray. 2014. Marine renewable energy, electromagnetic (EM) fields and EM-sensitive animals. Pp. 61–79 in Marine Renewable Energy Technology and Environmental Interactions. M.A. Shields, and A.I.L. Payne, eds, Springer, Dordrecht, the Netherlands, https://doi.org/10.1007/978-94-017-8002-5_6.
Gross, M.R., J. Repka, C.T. Robertson, D.H. Secor, and W. Van Winkle. 2002. Sturgeon conservation: Insights from elasticity analysis. Pp. 13–29 in Biology, Management, and Protection of North American Sturgeons, vol. 28. W. VanWinkle, P.J. Anders, D.H. Secor, and D.A. Dixon, eds, American Fisheries Society, Bethesda, MD, USA.
Hobson, V.J., D. Righton, J.D. Metcalfe, and G.C. Hays. 2007. Vertical movements of North Sea cod. Marine Ecology Progress Series 347:101–110, https://doi.org/10.3354/meps07047.
Hutchison, Z.L., A.B. Gill, P. Sigray, H. He, and J.W. King. 2020. Anthropogenic electromagnetic fields (EMF) influence the behaviour of bottom-dwelling marine species. Scientific Reports 10(1):4219, https://doi.org/10.1038/s41598-020-60793-x.
Kalmijn, A.J. 1971. The electric sense of sharks and rays. Journal of Experimental Biology 55(2):371–383.
Kalmijn, A.J. 1978. Experimental evidence of geomagnetic orientation in elasmobranch fishes. Pp. 347–353 in Animal Migration, Navigation, and Homing. Proceedings in Life Sciences, K. Schmidt-Koenig, and W. T. Keeton, eds, Springer Berlin Heidelberg, https://doi.org/10.1007/978-3-662-11147-5_34.
Kavet, R., M.T. Wyman, and A.P. Klimley. 2016. Modeling magnetic fields from a DC power cable buried beneath San Francisco Bay based on empirical measurements. PLoS ONE 11(2):e0148543, https://doi.org/10.1371/journal.pone.0148543.
Kempster, R.M., N.S. Hart, and S.P. Collin. 2013. Survival of the stillest: Predator avoidance in shark embryos. PLoS ONE 8(1):e52551, https://doi.org/10.1371/journal.pone.0052551.
Kilfoyle, A.K., R.F. Jermain, M.R. Dhanak, J.P. Huston, and R.E. Spieler. 2017. Effects of EMF emissions from undersea electric cables on coral reef fish. Bioelectromagnetics 9(10):22092, https://doi.org/10.1002/bem.22092.
Kimber, J.A., D.W. Sims, P.H. Bellamy, and A.B. Gill. 2011. The ability of a benthic elasmobranch to discriminate between biological and artificial electric fields. Marine Biology 158(1):1–8, https://doi.org/10.1007/s00227-010-1537-y.
Kimber, J.A., D.W. Sims, P.H. Bellamy, and A.B. Gill. 2014. Elasmobranch cognitive ability: Using electroreceptive foraging behaviour to demonstrate learning, habituation and memory in a benthic shark. Animal Cognition 17(1):55–65, https://doi.org/10.1007/s10071-013-0637-8.
Kirschvink, J.L. 1992. Uniform magnetic fields and double-wrapped coil systems: Improved techniques for the design of bioelectromagnetic experiments. Bioelectromagnetics 13(5):401–411, https://doi.org/10.1002/bem.2250130507.
Klimley, A.P. 1993. Highly directional swimming by scalloped hammerhead sharks, Sphyrna lewini, and subsurface irradiance, temperature, bathymetry, and geomagnetic field. Marine Biology 117(1):1–22, https://doi.org/10.1007/bf00346421.
Lohmann, K., N. Pentcheff, G. Nevitt, G. Stetten, R. Zimmer-Faust, H. Jarrard, and L. Boles. 1995. Magnetic orientation of spiny lobsters in the ocean: Experiments with undersea coil systems. Journal of Experimental Biology 198(Pt 10):2,041–2,048.
Love, M.S., M.M. Nishimoto, S. Clark, and A.S. Bull. 2015. Identical response of caged rock crabs (genera Metacarcinus and Cancer) to energized and unenergized undersea power cables in Southern California, USA. Bulletin of the Southern California Academy of Sciences 14(1):11.
Love, M.S., M.M. Nishimoto, S. Clark, M. McCrea, and A.S. Bull. 2017. Assessing potential impacts of energized submarine power cables on crab harvests. Continental Shelf Research 151:23–29, https://doi.org/10.1016/j.csr.2017.10.002.
Malagoli, D., M. Lusvardi, F. Gobba, and E. Ottaviani. 2004. 50 Hz magnetic fields activate mussel immunocyte p38 MAP kinase and induce HSP70 and 90. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 137(1):75–79, https://doi.org/10.1016/j.cca.2003.11.007.
Mapstone, B.D. 1995. Scalable decision rules for environmental impact studies: Effect size, type I, and type II Errors. Ecological Applications 5(2):401–410, https://doi.org/10.2307/1942031.
Nathan, R. 2008. An emerging movement ecology paradigm. Proceedings of the National Academy of Sciences of the United States of America 105(49):19,050–19,051, https://doi.org/10.1073/pnas.0808918105.
Newton, K.C., A.B. Gill, and S.M. Kajiura. 2019. Electroreception in marine fishes: Chondrichthyans. Journal of Fish Biology 95:135–154, https://doi.org/10.1111/jfb.14068.
Nichol, D.G., and D.A. Somerton. 2002. Diurnal vertical migration of the Atka mackerel Pleurogrammus monopterygius as shown by archival tags. Marine Ecology Progress Series 239:193–207, https://doi.org/10.3354/meps239193.
Nordmann, G.C., T. Hochstoeger, and D.A. Keays. 2017. Magnetoreception—A sense without a receptor. PLOS Biology 15(10):e2003234, https://doi.org/10.1371/journal.pbio.2003234.
Nyqvist, D., C. Durif, M. G. Johnsen, K. De Jong, T.N. Forland, and L.D. Sivle. 2020. Electric and magnetic senses in marine animals, and potential behavioral effects of electromagnetic surveys. Marine Environmental Research 155:104888, https://doi.org/10.1016/j.marenvres.2020.104888.
Overholtz, W.J., and K.D. Friedland. 2002. Recovery of the Gulf of Maine-Georges Bank Atlantic herring (Clupea harengus) complex: Perspectives based on bottom trawl survey data. Fishery Bulletin 100(3):593–608, https://spo.nmfs.noaa.gov/sites/default/files/pdf-content/2002/1003/overho.pdf.
Porsmoguer, S.B., D. Bănaru, C.F. Boudouresque, I. Dekeyser, and C. Almarcha. 2015. Hooks equipped with magnets can increase catches of blue shark (Prionace glauca) by longline fishery. Fisheries Research 172(1):345–351, https://doi.org/10.1016/j.fishres.2015.07.016.
Putman, N.F., K.J. Lohmann, E.M. Putman, T.P. Quinn, A.P. Klimley, and D.L.G. Noakes. 2013. Evidence for geomagnetic imprinting as a homing mechanism in Pacific salmon. Current Biology 23(4):312–316, https://doi.org/10.1016/j.cub.2012.12.041.
Putman, N.F., A.M. Meinke, and D.L. Noakes. 2014. Rearing in a distorted magnetic field disrupts the “map sense” of juvenile steelhead trout. Biology Letters 10(6):20140169, https://doi.org/10.1098/rsbl.2014.0169.
Rothermel, E.R., M.T. Balazik, J.E. Best, M.W. Breece, D.A. Fox, B.I. Gahagan, D.E. Haulsee, A.L. Higgs, M.H.P. O’Brien, M.J. Oliver, and others. 2020. Comparative migration ecology of striped bass and Atlantic sturgeon in the US Southern mid-Atlantic bight flyway. PLoS ONE 15(6):1–24, https://doi.org/10.1371/journal.pone.0234442.
Scott, K., P. Harsanyi, and A.R. Lyndon. 2018. Understanding the effects of electromagnetic field emissions from Marine Renewable Energy Devices (MREDs) on the commercially important edible crab, Cancer pagurus (L.). Marine Pollution Bulletin 131:580–588, https://doi.org/10.1016/j.marpolbul.2018.04.062.
Secor, D.H. 2015. Migration Ecology of Marine Fishes. Johns Hopkins University Press, Baltimore, MD, USA, 291 pp.
Secor, D.H., F. Zhang, M.H.P. O’Brien, and M. Li. 2019. Ocean destratification and fish evacuation caused by a Mid-Atlantic tropical storm. ICES Journal of Marine Science 76(2):573–584, https://doi.org/10.1093/icesjms/fsx241.
Sherwood, J., S. Chidgey, P. Crockett, D. Gwyther, P. Ho, S. Stewart, D. Strong, B. Whitely, and A. Williams. 2016. Installation and operational effects of a HVDC submarine cable in a continental shelf setting: Bass Strait, Australia. Journal of Ocean Engineering and Science 1(4):337–353, https://doi.org/10.1016/j.joes.2016.10.001.
Sisneros, J.A., T.C. Tricas, and C.A. Luer. 1998. Response properties and biological function of the skate electrosensory system during ontogeny. Journal of Comparative Physiology A 183(1):87–99, https://doi.org/10.1007/s003590050237.
Soares-Ramos, E.P.P., L. de Oliveira-Assis, R. Sarrias-Mena, and L.M. Fernández-Ramírez. 2020. Current status and future trends of offshore wind power in Europe. Energy 202:117787, https://doi.org/10.1016/j.energy.2020.117787.
Stankevičiūtė, M., M. Jakubowska, J. Pažusienė, T. Makaras, Z. Otremba, B. Urban-Malinga, D.P. Fey, M. Greszkiewicz, G. Sauliutė, J. Baršienė, and E. Andrulewicz. 2019. Genotoxic and cytotoxic effects of 50 Hz 1 mT electromagnetic field on larval rainbow trout (Oncorhynchus mykiss), Baltic clam (Limecola balthica) and common ragworm (Hediste diversicolor). Aquatic Toxicology 208:109–117, https://doi.org/10.1016/j.aquatox.2018.12.023.
Stoddard, P.K. 2010. Electrical signals. Pp. 601–610 in Encyclopedia of Animal Behavior. M.D. Breed and J. Moore, eds, Academic Press, Oxford, https://doi.org/10.1016/B978-0-08-045337-8.00002-4.
Taormina, B., J. Bald, A. Want, G. Thouzeau, M. Lejart, N. Desroy, and A. Carlier. 2018. A review of potential impacts of submarine power cables on the marine environment: Knowledge gaps, recommendations and future directions. Renewable and Sustainable Energy Reviews 96:380–391, https://doi.org/10.1016/j.rser.2018.07.026.
Taormina, B., C. Di Poi, A.L. Agnalt, A. Carlier, N. Desroy, R.H. Escobar-Lux, J.F. D’eu, F. Freytet, and C.M.F. Durif. 2020a. Impact of magnetic fields generated by AC/DC submarine power cables on the behavior of juvenile European lobster (Homarus gammarus). Aquatic Toxicology 220:105401, https://doi.org/10.1016/j.aquatox.2019.105401.
Taormina, B., M. Laurans, M.P. Marzloff, N. Dufournaud, M. Lejart, N. Desroy, D. Leroy, S. Martin, and A. Carlier. 2020b. Renewable energy homes for marine life: Habitat potential of a tidal energy project for benthic megafauna. Marine Environmental Research 161:105131, https://doi.org/10.1016/j.marenvres.2020.105131.
Tricas, T.C., and J.A. Sisneros. 2004. Ecological functions and adaptations of the elasmobranch electrosense. Pp. 308–329 in The Senses of Fish. G. von der Emde, J. Mogdans, and B.G. Kapoor, eds, Springer, Dordrecht, https://doi.org/10.1007/978-94-007-1060-3_14.
Walker, M.M., T.E. Dennis, and J.L. Kirschvink. 2002. The magnetic sense and its use in long-distance navigation by animals. Current Opinion in Neurobiology 12(6):735–744, https://doi.org/10.1016/S0959-4388(02)00389-6.
Walli, A., S.L.H. Teo, A. Boustany, C.J. Farwell, T. Williams, H. Dewar, E. Prince, and B.A. Block. 2009. Seasonal movements, aggregations and diving behavior of Atlantic bluefin tuna (Thunnus thynnus) revealed with archival tags. PLoS ONE 4(7), https://doi.org/10.1371/journal.pone.0006151.
Westerberg, H., and I. Lagenfelt. 2008. Sub-sea power cables and the migration behaviour of the European eel. Fisheries Management and Ecology 15(5–6):369–375, https://doi.org/10.1111/j.1365-2400.2008.00630.x.
WHO (World Health Organization). 2020. Electromagnetic fields, https://www.who.int/health-topics/electromagnetic-fields#tab=tab_1.
Wilding, T.A., A.B. Gill, A. Boon, E. Sheehan, J.C. Dauvin, J.-P.P. Pezy, F. O’Beirn, U. Janas, L. Rostin, I. De Mesel, and others. 2017. Turning off the DRIP (‘Data-rich, information-poor’)—Rationalising monitoring with a focus on marine renewable energy developments and the benthos. Renewable and Sustainable Energy Reviews 74:848–859, https://doi.org/10.1016/j.rser.2017.03.013.
Winemiller, K.O., and K.A. Rose. 1992. Patterns of life-history diversification in North American fishes: Implications for population regulation. Canadian Journal of Fisheries and Aquatic Sciences 49(10):2,196–2,218, https://doi.org/10.1139/f92-242.
Woodruff, D.L., I.R. Schultz, K.E. Marshall, J.A. Ward, and V.I. Cillinan. 2012. Effects of Electromagnetic Fields on Fish and Invertebrates. Task 2.1.3: Effects on Aquatic Organisms, Fiscal Year 2011 Progress Report. 68 pp.
Wyman, M.T., A. Peter Klimley, R.D. Battleson, T.V. Agosta, E.D. Chapman, P.J. Haverkamp, M.D. Pagel, and R. Kavet. 2018. Behavioral responses by migrating juvenile salmonids to a subsea high-voltage DC power cable. Marine Biology 165(8):134, https://doi.org/10.1007/s00227-018-3385-0.